A hidrák a ülő koelenterátumok egy speciális neme, amelyek megjelenésükben és életmódjukban a növényekre hasonlítanak, de mégis az állatvilágba tartoznak. Idegrendszer a hidra úgy van kialakítva, hogy biztosítsa a lény kinyerési képességét elegendő mennyiségbenétel.
Nem könnyű kitalálni, hogy a hidra milyen típusú idegrendszerrel rendelkezik, mivel ez a szerkezet meglehetősen egyszerű, és nemcsak ezekben a lényekben, hanem egyes medúzafajokban és más primitív állatokban is megtalálható. A hidrák viszonylag kisméretű állati szervezetek, amelyek mérete 2-20 mm.
Az idegrendszert alkotó sejtek csillag alakúak, melyeket sugarak kötnek össze egymással, idegi hálózatot alkotva. Az idegrendszer a bőr-izomsejtek alatt található. A hidrák nem rendelkeznek a külső vagy belső ingerek által okozott elektromos impulzusok központi érzékelésére szolgáló szervvel. Maximális összeg neuronok körülbelül 5000 db. és mind összefüggenek egymással.
A hidra idegrendszerét diffúz plexusnak nevezik, mivel van egy szétszórt és heterogén plexus. A diffúz plexus kondenzációja figyelhető meg a talp területén, szájüregés csápok. A legújabb tanulmányok kimutatták, hogy a száj területén egy ideggyűrű található, amelynek szerkezete hasonló a hydromedusa esernyő széle mentén elhelyezkedő ideggyűrűhöz.
A hidra idegrendszere rendkívül primitív, ezért az őt alkotó sejtek nem különülnek el egyértelműen motoros, interkaláris és szenzoros sejtekre. Ugyanakkor figyelembe kell venni, hogy ennek a lénynek az idegrendszerében még mindig létezik egy bizonyos sejtosztódás. Az idegsejteknek 2 fő típusa van - ganglion és érzékszervi.
Ennek a két sejttípusnak a szerkezete alapvető különbségeket mutat. Az érzékeny sejtek a hámrétegen keresztül helyezkednek el, és 1 mozdulatlan flagellumjuk van mikroszkopikus bolyhokkal tarkítva. Ez a flagellum jön ki külső környezetés kívülről ható ingereket vezet le. A ganglion típusú sejtek a hám-izomréteg legtövében helyezkednek el, így folyamataik nem tudják érzékelni a kívülről ható ingereket, de szükség esetén aktívan részt vesznek az izomösszehúzódásban.
Morfológiai összetételüket tekintve a hidra idegsejtek túlnyomó többsége bipoláris, ami jobb vezetőképességet biztosít számukra, és azt a képességet, hogy megfelelően reagáljanak a szervezet testét a külső környezetből érkező ingerekre.
A hidra idegrendszer primitív felépítése ellenére a vezetőképességet továbbra is nemcsak elektromos, hanem még mindig biztosítja kémiai reakciók. Az olyan szervezetben található kémiai neurotranszmitterek, mint a hidra, közé tartozik a szerotonin, dopamin, gamma-aminosav, noradrenalin, glutamát, glicin és ezen kívül még egy beteg mennyiség. különböző típusok neuropeptidek.
Mindezek vegyi anyagok inkább az összetett állati szervezetekre jellemzőek, de kis részük a protozoonokban is megtalálható. Annak ellenére, hogy a hidrának nincs központi idegrendszere, mégis képes érzékelni a fényingereket. Viszonylag a közelmúltban még az olyan organizmusokat is teljesen képtelennek tartották megkülönböztetni, mint a medúza, de később olyan speciális sejteket fedeztek fel, amelyek lehetővé teszik, hogy ezek az óceánon át sodródó lények különbséget tegyenek a fény és a sötét között, és megválasszák a mozgás irányát. Ez rendkívül hatékony, mert több felületi rétegek víz él nagy mennyiség kis rákfélék és más szervezetek, amelyekkel a medúza táplálkozik.
A Hydra hasonló mechanizmussal rendelkezik a fény és a sötétség felismerésére. Egy speciális érzékeny fehérje, más néven opszin segít a hidráknak a fény felismerésében. A hidra testéből kivont fehérje genetikai elemzése számos hasonlóságot tárt fel egy, az emberben található hasonló fehérjével. Egy hasonló tanulmány kimutatta, hogy az emberekben és a hidrában található opszin fehérje közös eredetű.
A hidra idegrendszere meglehetősen hatékony, és biztosítja ennek a lénynek a legjobb feltételeket a túléléshez. A hidra testével való minimális érintkezés esetén az izgalom, amely testének egy pontjáról ered, gyorsan átterjed a többire is. Tekintve, hogy ingerület azonnal elterjed a hidra testében, a bőr-izomrendszer gyors összehúzódása figyelhető meg, aminek következtében a lény egész teste gyorsan lerövidül. A külső ingerre adott ilyen válasz feltétel nélküli reflexnek minősül.
Az idegsejtek a hidratest más szöveteihez hasonlóan jelentős regenerációs képességgel rendelkeznek. Ha a hidra több részre oszlik, ezek a felek később válhatnak független szervezetés újratermesztjük az elveszett részeket.
Annak ellenére, hogy a hidrák általában hosszú ideig egy helyen maradnak, ha szükséges, ez a lény lassan mozoghat, hogy kényelmesebb helyet találjon a zsákmányra. A hidra mozgásának sajátosságait is nagymértékben meghatározza ennek a lénynek az idegrendszerének felépítése.
Hidra mozgások. Az ektoderma hám-izomsejtjei olyan rostokkal rendelkeznek, amelyek összehúzódhatnak. Ha egyidejűleg összehúzódnak, a hidra egész teste megrövidül. Ha az egyik oldalon csökken a bürokrácia a sejtekben, akkor a hidra ebbe az irányba billen. Ezen rostok munkájának köszönhetően a hidra csápjai megmozdulnak, és az egész teste megmozdul (13.4. ábra).
Reakciók a Hydra irritációra. Az ektodermában található idegsejteknek köszönhetően a hidra érzékeli külső irritációk: fény, tapintás, némi vegyszer. Ezeknek a sejteknek a folyamatai összezáródnak, hálót alkotva. Így jön létre a szerkezetében legegyszerűbb idegrendszer, ún diffúz (13.5. ábra). Az idegsejtek többsége a talp közelében és a csápokon található. Az idegrendszer és a hám izomsejtek munkájának megnyilvánulása a feltétel nélküli hidra reflex - az érintésre válaszul a csápok meghajlítása.
| Rizs. 13.4. Hidra mozgás diagram |
 |
| Rizs. 13.5. Hidra idegrendszer |
A külső réteg csípősejteket is tartalmaz, amelyek kapszulákat tartalmaznak egy csavart vékony csővel - a szúrófonal. Egy érzékeny szőr kilóg a sejtből. Elég enyhén megérinteni, és a cérna kilökődik a kapszulából, és átszúrja az ellenség vagy a zsákmány testét. A szúrószálon keresztül méreg jut hozzá, és az állat elpusztul. A legtöbb csípősejt a csápokban található.
Hidraregeneráció. A kicsi, kerek, közbenső ektoderma sejtek képesek átalakulni más típusú sejtekké. Szaporodásuknak köszönhetően a hidra gyorsan újjáépíti a sérült testrészt. Elképesztő az állat regenerálódásának képessége: amikor a hidrát 200 részre osztották, mindegyikből egy egész állatot sikerült helyreállítani!
Hidra táplálkozás. Az endodermiszben található mirigysejtekés flagellákkal felszerelve emésztő sejtek. A mirigysejteket ellátják bélüreg emésztőnedveknek nevezett anyagok. Ezek az anyagok elpusztítják a zsákmányt, mikroszkopikus darabokra bontják. A flagellák segítségével az emésztősejtek maguk felé tolják és elfogják őket, pszeudopodiákat képezve. Belső üreg Nem véletlenül nevezik a hidrát bélnek: benne kezdődik meg a táplálék emésztése. De a táplálék végül lebomlik az emésztősejtek emésztőüregeiben. Az emésztetlen táplálékmaradványokat a szájon keresztül távolítják el a bélüregből.
Kiválasztás káros anyagok a hidra élete során keletkezik, az ektodermán keresztül kerül a vízbe
Sejtkölcsönhatás. A hidrasejtek közül csak az emésztősejtek emésztik meg a táplálékot, de nem csak önmaguknak, hanem minden más sejtnek is tápanyagot biztosítanak. Viszont a „szomszédok” alkotnak legjobb körülmények közöttélet a tápanyag-beszállítók számára. Gondoljunk csak a hidravadászatra – most elmagyarázhatja, hogy az ideg-, szúró-, hám-izom- és mirigysejtek összehangolt munkája hogyan biztosítja az emésztősejtek működését. És ezek a sejtek megosztják munkájuk eredményét szomszédaikkal. Anyag az oldalról
Hogyan szaporodik a hidra? Az ivartalan szaporodás során a köztes sejtek osztódása következtében rügy képződik. A bimbó nő, csápok jelennek meg rajta, és száj tör ki közöttük. A másik végén a talp formálódik. A kis hidra elválik az anya testétől, lesüllyed a fenékre, és önálló életet kezd.
A hidra szexuálisan is szaporodik. A hidra egy hermafrodita: ektodermájának egyes kiemelkedései közbenső sejtekből, másokban tojásokból képződnek a spermiumok. Miután elhagyta a hidra testét, a spermiumok követik a vizet más egyedekhez. Miután megtalálták a petéket, megtermékenyítik őket. Zigóta képződik, amely körül sűrű membrán jelenik meg. Ez a megtermékenyített tojás a hidra testében marad. Általában szexuális szaporodásősszel történik. Télen a kifejlett hidrák elpusztulnak, és a tojások a tározó alján élik túl a telet. Tavasszal a zigóta osztódni kezd, és két sejtréteget képez. Belőlük egy kis hidra fejlődik.
Ezen az oldalon a következő témákban található anyagok:
A szivacsszaporodás állapota
Irritáció és mozgásbiológiai jelentés
A hidratest sejtjeinek felépítésének és működésének sajátosságai
A hidra életfolyamatainak jellemzői
Hasonlítsa össze a hidraszúró sejt szerkezetét és a csalánlevél bőrét!
Kérdések ezzel az anyaggal kapcsolatban:
-
A hidra teste úgy néz ki, mint egy hosszúkás zsák, amelynek falai két sejtrétegből állnak - ektodermaÉs endoderma.
Közöttük egy vékony, zselatinos, nem sejtes réteg található - mesoglea, támaszként szolgál.

Az ektoderma képezi az állat testének borítását, és többféle sejtből áll: hám-izmos, közbülsőÉs szúrós.
Közülük a legtöbb hám-izmos.
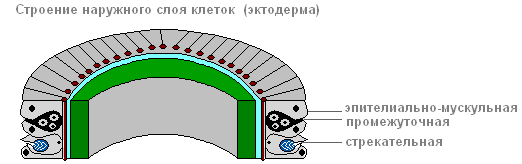
Ektoderm

hám izomsejt
Következtében izomrostok, minden sejt tövében fekszik, a hidra teste összehúzódhat, megnyúlhat és meghajolhat.
A hám-izomsejtek között kisméretű, kerek, nagy sejtmaggal és kis mennyiségű citoplazmával rendelkező sejtcsoportok találhatók, ún. közbülső.

Amikor a hidra teste megsérül, elkezdenek gyorsan növekedni és osztódni. Más típusú sejtekké alakulhatnak át a hidratestben, kivéve a hám-izmos sejteket.
Az ektoderma tartalmaz szúró sejtek, támadásra és védekezésre szolgál. Főleg a hidra csápjain helyezkednek el. Minden szúrósejt tartalmaz egy ovális kapszulát, amelyben a szúrószál feltekercselődik.
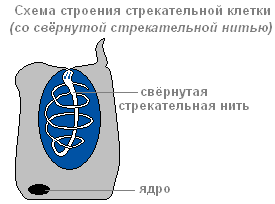
Feltekeredett szúrószálú szúrósejt felépítése
Ha a zsákmány vagy az ellenség megérinti a szúró sejten kívül található érzékeny szőrszálat, az irritáció hatására a szúrószál kilökődik és átszúrja az áldozat testét.
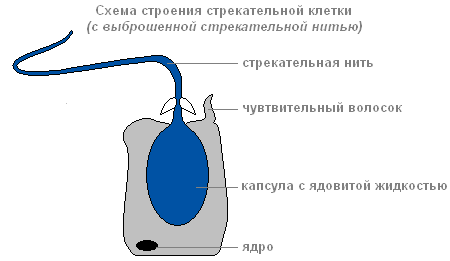
A csípősejt felépítése kiselejtezett szúrószállal
A menetcsatornán keresztül olyan anyag kerül az áldozat testébe, amely megbéníthatja az áldozatot.
A szúrósejteknek többféle típusa van. A szálak valami átszúrják bőrállatokat és mérget fecskendeznek a szervezetükbe. Mások szálai a zsákmány köré tekernek. A harmadik szálai nagyon ragadósak és az áldozathoz tapadnak. Általában a hidra több csípős sejtet „lelő”. A lövés után a szúró sejt elhal. Új szúrósejtek keletkeznek abból közbülső.
A sejtek belső rétegének szerkezete
Az endoderma belülről kibéleli a teljes bélüreget. Magába foglalja emésztő-izmosÉs mirigyes sejteket.

Endoderm
Emésztőrendszer
Több emésztőizomsejt van, mint mások. Izomrostok redukcióra képesek. Amikor lerövidülnek, a hidra teste elvékonyodik. Az összetett mozgások (mozgás „buktatással”) az ektoderma és endoderma sejtek izomrostjainak összehúzódása miatt következnek be.
Az endoderma emésztőizomsejtjei mindegyike 1-3 flagellával rendelkezik. Habozó flagella vízáramot hoznak létre, amely az élelmiszer-részecskéket a sejtek felé hajtja. Az endoderma emésztőizomsejtjei képesek kialakulni állábúak, felfogja és megemészti a kis élelmiszer-részecskéket az emésztőüregekben.
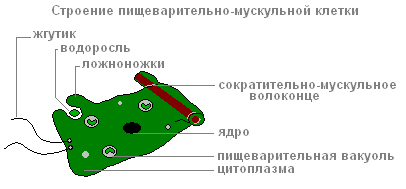
Az emésztőizomsejt szerkezete
Az endodermában lévő mirigysejtek a bélüregbe szekretálódnak emésztőnedv, amely cseppfolyósítja és részben megemészti az ételt.

A mirigysejt felépítése
A zsákmányt a csápok csípősejtek segítségével fogják be, amelyek mérge gyorsan megbénítja a kis áldozatokat. A csápok összehangolt mozgásával a zsákmányt a szájhoz viszik, majd a testösszehúzódások segítségével a hidrát „ráhelyezik” az áldozatra. Az emésztés a bélüregben kezdődik ( üreges emésztés), a hám-izom endoderma sejtek emésztő vakuólumaiban végződik ( intracelluláris emésztés). Tápanyagok eloszlik a hidra testében.
Amikor az emésztőüreg a zsákmány nem emészthető maradványait és a sejtanyagcsere hulladékát tartalmazza, összehúzódik és kiürül.
Lehelet
A hidra vízben oldott oxigént lélegez be. Nincsenek légzőszervei, és testének teljes felületén szívja fel az oxigént.
Keringési rendszer
Hiányzó.
Kiválasztás
Kiválasztás szén-dioxidés az életfolyamat során keletkező egyéb felesleges anyagokat a külső réteg sejtjeiből közvetlenül a vízbe, a belső réteg sejtjeiből pedig a bélüregbe juttatják, majd ki.
Idegrendszer
A bőr és az izomsejtek alatt sejtek találhatók csillag alakú. Ez idegsejtek(1). Összekapcsolódnak egymással, és ideghálózatot alkotnak (2).

Az idegrendszer és a hidra ingerlékenysége
Ha megérinti a hidrát (2), akkor az idegsejtekben gerjesztés (elektromos impulzusok) lép fel, amely azonnal szétterjed a teljes ideghálózatban (3) és a bőr-izomsejtek összehúzódását okozza, és a hidra egész teste lerövidül ( 4). A hidra testének válasza az ilyen irritációra az feltétlen reflex.
Nemi sejtek
Az őszi hideg időjárás közeledtével a hidra ektodermájában lévő köztes sejtekből csírasejtek képződnek.
A csírasejteknek két típusa van: a tojás vagy a női csírasejtek és a hímivarsejtek, vagyis a hímivarsejtek.
A peték közelebb helyezkednek el a hidra alapjához, a spermiumok a szájhoz közelebb elhelyezkedő gumókban fejlődnek ki.
petesejt A hidra hasonló az amőbához. Pszeudopodákkal van felszerelve, és gyorsan növekszik, felszívja a szomszédos köztes sejteket.

A hidra petesejtek szerkezete

A hidra sperma szerkezete
SpermaÁltal kinézet lobogó protozoákra hasonlítanak. Elhagyják a hidra testét, és hosszú flagellum segítségével úsznak.
Megtermékenyítés. Reprodukció
A spermium a petesejttel együtt felúszik a hidrához, és behatol annak belsejébe, és mindkét nemi sejt magja egyesül. Ezt követően a pszeudopodák visszahúzódnak, a sejt lekerekedik, felületén vastag héj szabadul fel - tojás képződik. Amikor a hidra meghal és megsemmisül, a tojás életben marad és a fenékre esik. A meleg idő beköszöntével élő sejt, amely a védőhéjon belül helyezkedik el, osztódni kezd, a keletkező sejtek két rétegben helyezkednek el. Belőlük egy kis hidra fejlődik ki, amely a tojáshéj törésén keresztül jön ki. Így a többsejtű állati hidra élete kezdetén csak egy sejtből áll - egy tojásból. Ez arra utal, hogy a Hydra ősei egysejtű állatok voltak.
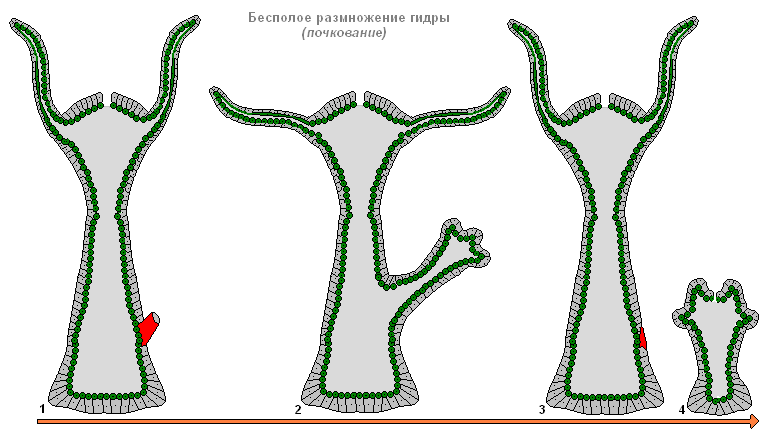
Aszexuális szaporodás hidra
Nál nél kedvező feltételek A hidra ivartalanul szaporodik. Az állat testén (általában benne alsó harmadát test) rügy keletkezik, megnő, majd csápok alakulnak ki, és áttörik a száj. Az anya testéből származó fiatal hidrarügyek (ebben az esetben az anya- és lánypolipok csápokkal vannak rögzítve az aljzathoz, és különböző irányokba húzódnak) és elvezetik független képélet. Ősszel a hidra ivaros szaporodásnak indul. A testen, az ektodermában ivarmirigyek képződnek - nemi mirigyek, és bennük a csírasejtek köztes sejtekből fejlődnek ki. Amikor a hidra ivarmirigyek kialakulnak, medusoid csomó képződik. Ez azt sugallja, hogy a hidra ivarmirigyek erősen leegyszerűsített sporiferek, az elveszett meduzoid nemzedék szervvé történő átalakulásának sorozatának utolsó szakasza. A legtöbb hidrafaj kétlaki, kevésbé gyakori a hermafroditizmus. A hidrapeték gyorsan növekednek a környező sejtek fagocitizálásával. Az érett tojások átmérője eléri a 0,5-1 mm-t. A megtermékenyítés a hidra testében történik: az ivarmirigyben lévő speciális lyukon keresztül a spermium behatol a petesejtbe, és összeolvad vele. A zigóta teljes egyenletes fragmentáción megy keresztül, melynek eredményeként coeloblastula képződik. Ezután vegyes delamináció (bevándorlás és delamináció kombinációja) eredményeként gastruláció lép fel. Az embrió körül sűrű védőburok (embriotheca) képződik, gerincszerű kinövésekkel. A gastrula szakaszban az embriók felfüggesztett animációba lépnek. A kifejlett hidrák elpusztulnak, az embriók a fenékre süllyednek és áttelelnek. Tavasszal folytatódik a fejlődés, az endoderma parenchymájában a sejtek divergenciájával bélüreg alakul ki, majd kialakulnak a csápok rudimentumai, a héj alól egy fiatal hidra bújik elő. Így a legtöbb tengeri hidroiddal ellentétben a hidrának nincsenek szabadon úszó lárvái, és fejlődése közvetlen.
Regeneráció
A Hydra nagyon magas regenerációs képességgel rendelkezik. Ha keresztben több részre vágják, mindegyik rész visszaállítja a „fejet” és a „lábat”, megtartva az eredeti polaritást - a száj és a csápok azon az oldalon fejlődnek ki, amely közelebb volt a test orális végéhez, a szár és a talp pedig ezen. a töredék aborális oldala. Az egész szervezet helyreállítható egyedi kis testdarabokból (kevesebb, mint 1/100 térfogat), csápdarabokból, valamint sejtszuszpenzióból. Ugyanakkor magát a regenerációs folyamatot nem kíséri növekedés sejtosztódásés a morfhallaxis tipikus példája.
Mozgalom
BAN BEN nyugodt állapot a csápok több centiméterre nyúlnak ki. Az állat lassan mozgatja őket egyik oldalról a másikra, prédára lesve. Ha szükséges, a hidra lassan mozoghat.
"Gyalogos" közlekedési mód

A hidra "séta" mozgási módja
Miután a testét (1) meggörbítette, és csápjait egy tárgy (szubsztrátum) felületéhez rögzítette, a hidra a talpat (2) a test elülső végéhez húzza. Ezután a hidra sétáló mozgása megismétlődik (3,4).
"Buktatós" mozgásmód

A hidra "buktatós" mozgási módja
Egy másik esetben úgy tűnik, hogy a feje fölött bukdácsol, és csápjaival és talpával felváltva tapad a tárgyakhoz (1-5).
ábra: Édesvízi hidra szerkezete. A hidra radiális szimmetriája
Az édesvízi hidrapolip élőhelye, szerkezeti jellemzői és életfunkciói
A tiszta, átlátszó vízzel rendelkező tavakban, folyókban vagy tavakban a vízinövények szárán egy kis áttetsző állat található - polip hidra(a „polip” jelentése „többlábú”). Ez egy kötődő vagy ülő, coelenterate állat, számos csápok. Egy közönséges hidra teste szinte szabályos hengeres alakú. Az egyik végén van száj 5-12 vékony, hosszú csápból álló corolla veszi körül, a másik vége szár formájában megnyúlt. egyetlen a végén. A talp segítségével a hidra különféle víz alatti tárgyakhoz rögzíthető. A hidra teste a szárral együtt általában legfeljebb 7 mm hosszú, de a csápok több centiméterre is kinyúlhatnak.
A hidra radiális szimmetriája
Ha egy képzeletbeli tengelyt rajzol a hidra teste mentén, akkor a csápjai minden irányban eltérnek ettől a tengelytől, mint a fényforrásból érkező sugarak. Lóg néhányról vízi növény, a hidra folyamatosan imbolyog és lassan mozgatja csápjait, lesben áll a prédára. Mivel a zsákmány bármely irányból megjelenhet, a sugárirányban elhelyezett csápok felelnek meg a legjobban ennek a vadászati módnak.
A sugárzási szimmetria általában a kötődő életmódot folytató állatokra jellemző.Hidra bélüreg
A hidra teste zsák alakú, amelynek falai két sejtrétegből állnak - a külső (ektoderma) és a belső (endoderma). A hidra testében van bélüreg(innen a típus neve - coelenterates).
A hidrasejtek külső rétege az ektoderma.

ábra: a sejtek külső rétegének felépítése - hidra ektoderma
A hidrasejtek külső rétegét - ektoderma. Mikroszkóp alatt a hidra külső rétegében - az ektodermában - többféle sejt látható. Itt leginkább bőrizmok vannak. Ezek a sejtek oldaluk érintésével létrehozzák a hidra fedelét. Minden ilyen sejt tövében összehúzódó izomrost található, amely fontos szerepet játszik az állat mozgásában. Amikor mindenki szálkás bőr-izmos a sejtek összehúzódnak, a hidra teste összehúzódik. Ha a rostok csak a test egyik oldalán húzódnak össze, akkor a hidra ebbe az irányba hajlik. Az izomrostok munkájának köszönhetően a hidra lassan mozoghat egyik helyről a másikra, felváltva „lépkedhet” a talpával és a csápjaival. Ez a mozdulat a fejed feletti lassú bukfencezéshez hasonlítható.
A külső réteg tartalmaz és idegsejtek. Csillag alakúak, mivel hosszú folyamatokkal vannak felszerelve.
A szomszédos idegsejtek folyamatai érintkeznek egymással és kialakulnak idegfonat, amely a hidra egész testét lefedi. Néhány folyamat megközelíti a bőr-izomsejteket.Hidra ingerlékenység és reflexek
A Hydra képes érzékelni az érintést, a hőmérséklet-változásokat, a vízben oldott különféle anyagok megjelenését és egyéb irritációkat. Emiatt idegsejtjei izgatottak lesznek. Ha egy vékony tűvel megérinti a hidrát, akkor az egyik idegsejt irritációjából származó izgalom a folyamatok mentén átkerül a többi idegsejtekbe, és azokból a bőr-izomsejtekbe. Emiatt az izomrostok összehúzódnak, és a hidra labdává zsugorodik.

Kép: Hydra ingerlékenysége
Ebben a példában egy összetett jelenséggel ismerkedünk meg az állati testben - reflex. A reflex három egymást követő szakaszból áll: az irritáció észlelése, gerjesztés átadása ettől az irritációtól az idegsejtek mentén és válasz testet bármilyen cselekvéssel. A hidra felépítésének egyszerűsége miatt reflexei nagyon egységesek. A jövőben sokkal összetettebb reflexekkel ismerkedünk meg jobban szervezett állatoknál.
Hidra szúró sejtek

Minta: hidraszál vagy csalánsejtek
A hidra egész teste és különösen a csápjai nagy számmal ülnek szúrós, vagy csalán sejteket. Mindegyik sejt rendelkezik összetett szerkezet. A citoplazmán és a sejtmagon kívül egy buborékszerű szúrókapszulát is tartalmaz, amibe egy vékony cső van összehajtva - szúró szál. Kilóg a ketrecből érzékeny haj. Amint egy rákféle, kis hal vagy más kis állat hozzáér egy érzékeny szőrhöz, a csípős fonal gyorsan kiegyenesedik, a vége kidobódik és átszúrja az áldozatot. A cérna belsejében áthaladó csatornán keresztül méreg kerül a zsákmány testébe a csípős kapszulából, kis állatok pusztulását okozva. Általában sok szúró sejtet lőnek ki egyszerre. Ezután a hidra a csápjaival a szájához húzza a zsákmányt és lenyeli. A csípős sejtek védelmet nyújtanak a hidra számára is. A halak és a vízi rovarok nem esznek hidrákat, amelyek megégetik ellenségeiket. A kapszulákból származó méreg a nagytestű állatok szervezetére gyakorolt hatásában csalánméregre emlékeztet.
A sejtek belső rétege a hidra endoderma

ábra: a sejtek belső rétegének felépítése - hidra endoderma
A sejtek belső rétege - endoderma A. A belső réteg - az endoderma - sejtjei összehúzódó izomrostokkal rendelkeznek, de ezeknek a sejteknek a fő szerepe a táplálék megemésztése. Emésztőnedvet választanak ki a bélüregbe, melynek hatására a hidra zsákmánya meglágyul és felbomlik. finom részecskék. A belső réteg egyes sejtjei több hosszú flagellával vannak felszerelve (mint a flagellált protozoákban). Flagella bent van állandó mozgásés a részecskéket a sejtek felé gereblyézi. A belső réteg sejtjei képesek állábúakat (mint egy amőba) kiszabadítani, és táplálékot fogni velük. A további emésztés a sejt belsejében, vakuólumokban (mint a protozoákban) megy végbe. Az emésztetlen ételmaradványok a szájon keresztül távoznak.
A hidrának nincsenek speciális légzőszervei, a vízben oldott oxigén a teste teljes felületén keresztül behatol a hidrába.Hidratáló regeneráció
A hidra testének külső rétege nagyon kicsi, kerek sejteket is tartalmaz, nagy magokkal. Ezeket a sejteket ún közbülső. Nagyon fontos szerepet játszanak a hidra életében. A test bármilyen károsodása esetén a sebek közelében található köztes sejtek gyorsan növekedni kezdenek. Belőlük bőr-izom, ideg- és egyéb sejtek képződnek, és a sebzett terület gyorsan begyógyul.
Ha keresztbe vágunk egy hidrát, akkor az egyik felén csápok nőnek, és megjelenik a száj, a másikon pedig egy szár. Kapsz két hidrát.
Az elveszett vagy sérült testrészek helyreállításának folyamatát ún regeneráció. A hidrának nagyon fejlett regenerációs képessége van.
A regeneráció ilyen vagy olyan mértékben más állatokra és emberekre is jellemző. Így a gilisztában egy egész szervezet regenerálható részeikből a kétéltűeknél (békák, gőték) egész végtagokat, a szem különböző részeit, farkát és belső szervek. Amikor egy személyt megvágnak, a bőr helyreáll.Hidra szaporodás
A hidra ivartalan szaporodása bimbózással

ábra: Hidra ivartalan szaporodás bimbózás útján
A hidra ivartalanul és ivarosan szaporodik. Nyáron egy kis gumó jelenik meg a hidra testén - a test falának kiemelkedése. Ez a gumó nő és nyúlik. A végén csápok jelennek meg, és egy száj tör ki közöttük. Így alakul ki a fiatal hidra, amely eleinte egy szár segítségével marad kapcsolatban az anyával. Kívülről mindez egy növényi hajtás bimbóból való kifejlődésére hasonlít (innen a jelenség neve - bimbózó). Amikor a kis hidra felnő, elválik az anya testétől, és önálló életet kezd.
Hidra ivaros szaporodás
Őszre, a kedvezőtlen körülmények beálltával a hidrák elpusztulnak, de előtte nemi sejtek fejlődnek ki szervezetükben. A csírasejteknek két típusa van: tojásdad, vagy nőstény, és spermiumok, vagy férfi reproduktív sejtek. A spermiumok hasonlóak a flagellated protozoákhoz. Elhagyják a hidra testét, és hosszú flagellum segítségével úsznak.

ábra: Hidra ivaros szaporodás
A hidra tojássejtje hasonló az amőbához, és állábúakkal rendelkezik. A spermium a petesejttel együtt felúszik a hidrához, és behatol annak belsejébe, és mindkét nemi sejt magja egyesül. Esemény megtermékenyítés. Ezt követően a pszeudopodákat visszahúzzuk, a sejtet lekerekítjük, és a felületén vastag héj képződik - a tojás. Ősz végén a hidra elpusztul, de a tojás életben marad, és a fenékre esik. Tavasszal a megtermékenyített tojás osztódni kezd, a kapott sejtek két rétegben vannak elrendezve. Belőlük egy kis hidra fejlődik ki, amely a meleg idő beálltával a tojáshéj törésén keresztül jön ki.
Így a többsejtű állati hidra élete elején egy sejtből áll - egy tojásból.CM. Nikitina, I.A. Vakoljuk (Kalinyingrád Állami Egyetem)
Hogyan működnek a hormonok a legfontosabb szabályozók valamint az anyagcsere és a szervezet különböző funkcióinak integrálói lehetetlen a specifikus jel vételére és végső átalakításra szolgáló rendszerek megléte nélkül. jótékony hatása, vagyis hormonkompetens rendszer nélkül. Más szavakkal, az exogén vegyületekre szervezeti szintű reakció jelenléte lehetetlen anélkül, hogy ezeknek a vegyületeknek a citorecepciója jelen lenne, és ennek megfelelően ezekben az állatokban nem léteznének olyan endogén vegyületek, amelyek rokonok azokkal, amelyekkel mi hatnak. Ez nem mond ellent az univerzális blokkok koncepciójának, amikor az alapvető molekuláris szerkezetek bekerülnek funkcionális rendszerekélő szervezetek szinte teljes halmazban megtalálhatók már az evolúció legkorábbi szakaszaiban, amelyek csak tanulmányozhatók, korlátozott számú molekulával képviselik és ugyanazt végzik. elemi függvények nemcsak egy királyság képviselői között, például ben különböző csoportok emlősök vagy akár különböző típusok, hanem különféle birodalmak képviselőiben is, beleértve a többsejtű és egysejtű szervezeteket, a magasabb rendű eukarióták és prokarióták.
Meg kell azonban jegyezni, hogy a gerincesekben hormonként működő vegyületek összetételére és funkcióira vonatkozó adatok a meglehetősen alacsony filogenetikai szintű taxonok képviselőiben csak most kezdenek megjelenni. Az alacsony filogenetikai szintű állatcsoportok közül a hidra, mint a koelenterátumok képviselője, a legprimitívebb, valódi idegrendszerrel rendelkező szervezet. A neuronok morfológiailag, kémiailag és valószínűleg funkcionálisan különböznek egymástól. Mindegyik neuroszekréciós granulátumot tartalmaz. A Hydra-ban a neuronális fenotípusok jelentős sokféleségét állapították meg. A hiposztómában 6-11 szinaptikusan kapcsolódó sejtből álló rendezett csoportok találhatók, amelyek a hidrák primitív ideg ganglionjainak bizonyítékának tekinthetők. Amellett, hogy viselkedési reakciókat biztosít, a hidra idegrendszer endokrin szabályozó rendszerként is szolgál, szabályozva az anyagcserét, a szaporodást és a fejlődést. A hidrákban az idegsejtek differenciálódnak a bennük lévő neuropeptidek összetétele szerint). Feltételezhető, hogy az oxitocin, a vazopresszin, a nemi szteroidok és a glükokortikoidok molekulái univerzálisak. A coelenterátusok képviselőiben is megtalálhatók. A fej és a talp aktivátorait (és inhibitorait) a hidratest metanolos kivonataiból izolálják. A tengeri kökörcsinekből izolált fejaktivátor összetételében és tulajdonságaiban hasonló a tehenek, patkányok, sertések, emberek hipotalamuszában és beleiben, valamint az utóbbiak vérében található neuropeptidhez. Emellett kimutatták, hogy gerincteleneknél és gerinceseknél is a ciklikus nukleotidok részt vesznek a sejtek neurohormonokra adott válaszának biztosításában, vagyis ezeknek az anyagoknak a hatásmechanizmusa két filogenetikailag eltérő vonalban azonos.
Célja ez a tanulmány A fentiek figyelembevételével az exogén biológiailag aktív (hormonális) vegyületek édesvízi hidrára gyakorolt komplex hatásának vizsgálatát választottuk.
Anyag és kutatási módszerek
A kísérlethez szükséges állatokat 1985. június-júliusban gyűjtötték össze. kórházban (a Nemonin folyó csatornája, Matrosovo falu, Polesie járás). A tartalomhoz való alkalmazkodás laboratóriumi körülmények- 10-14 nap. Anyagtérfogat: típus - Coelenterata; osztály - Hydrozoa; fajok - Hydra oligactis Pallas; mennyiség - 840. Az állatok számát a kísérlet elején tükrözik, és a szám növekedését nem vesszük figyelembe.
A munkához vízben oldódó anyagot használtak hormonális vegyületek oxitocin sorozat, az agyalapi mirigy elülső lebenye 1 ml (ip) kezdeti aktivitással (hifotocin - 5 egység, pituitrin - 5 egység, mammophysin - 3 egység, prefizon - 25 egység, gonadotropin - 75 egység) és egy szteroid - prednizolon - 30 mg, amely gerinceseknél háromrészes endokrin szabályozást biztosít, beleértve a hipotalamusz-hipofízis komplexet és a hámmirigyeket.
Az előkísérletekben az állattartó környezet 0,00002 és 20 ml ip/l közötti gyógyszerkoncentrációit alkalmaztuk.
Három tanulmányi csoport volt:
1. - a „+” vagy „-” reakció meghatározása minden általunk elfogadott koncentrációban;
2. - a koncentráció-tartomány meghatározása, amely biztosítja a krónikus üzemmódban végzett munkát különböző időtartamú;
3. - krónikus kísérlet.
A kísérlet figyelembe vette a Hydra bimbózó aktivitását. A kapott adatokat standard statisztikai feldolgozásnak vetették alá.
Kutatási eredmények
A hidrák „±” reakciójának meghatározásakor a vegyületek széles koncentráció-tartományában hármat választottunk ki (0,1 ml IP/L táptalaj, 0,02 ml IP/L táptalaj és 0,004 ml IP/L táptalaj).
A hidrák kontrollcsoportjában a bimbózás 0,0-0,4 rügy/hidra (Pa) szinten maradt 5 napig. A környezetben minimális koncentráció a prefizonnál 2,2 egyed/hidra, a pituitrinnél - 1,9 egyed/hidra volt a növekedés (a kontrollhoz viszonyított különbségek jelentősége rendkívül magas - 0,01 szignifikancia szint mellett). Közepes koncentrációban a hyfotocin, a mammophysin és a prefisone jól teljesített (1,8-1,9 egyed/hidra). A prednizolon minimális, és főleg átlagos koncentrációban 1,1-1,3 egyed/hidra szám növekedést okozott, ami jelentősen meghaladja a kontrollt.
A következő kísérletben csak optimális koncentrációjú hormonvegyületeket használtunk. A kísérlet időtartama 9 nap volt. A kísérlet kezdetén a kontroll és a kísérleti csoport nem volt megbízhatóan megkülönböztetve a Pa érték alapján. Kilenc napos kísérlet után a Pa értékek szignifikánsan különböztek a kísérleti csoportokban és a kontrollban 0,05-ös szignifikanciaszinttel (1. táblázat).
Asztal 1
A hormonális gyógyszerek hatása a hidra bimbózásra (Ra) és különbségeik jelentőségének valószínűsége (p)
szerda Ra változás R 1 nap 9 nap Ra 1 nap 9 nap Ellenőrzés 1,2±0,8 1,5±0,9 0,3±0,1 - - Gonadotropin 2,1±1,2 5,1±0,3 3,0±0,8 0,71 0,95 Prefison 1,1±0,7 4,9±2,0 3,8±1,3 0,13 0,97 Hyfotocin 1,8±0,8 6,1±2,2 4,3±1,4 0,58 0,99 Pituitrin 0,8±0,5 4,5±2,0 3,7±1,5 0,47 0,98 Mammophysin 1,1±0,3 5,3±2,0 4,2±1,7 0,15 0,99 Prednizolon 1,5±0,4 7,1±2,2 5,6±1,8 0,43 0,99 Ahogy a táblázatból is látszik, legmagasabb érték Az Ra-t akkor kapták, amikor az állatokat prednizolonban tartották. Minden peptidkészítmény megközelítőleg hasonló Pa-értéket ad (átlag 3,8 ± 0,5). Azonban itt is van eltérés. A legjobb hatás(4,3±1,4) akkor érhető el, ha az állatokat a neurohypophysis tisztított kivonatával - hyphotocinnal - tartalmazó környezetben tartják. Hatás szempontjából közel áll hozzá a mammophysin. A pituitrinnel és prefizonnal végzett kísérleti csoportokban az Ra értéke 3,7±1,5 és 3,8±1,3 volt. A legkisebb hatást a hidra gonadotropinnal történő befolyásolása éri el. Megbízhatatlan különbségek Ra-ban a hidra hormonális gyógyszerek oldataiba való behelyezése utáni első nap végén jelentkeznek. A kísérlet kilenc napja alatt a kontrollban Ra nem változott. A harmadik naptól kezdődően az összes kísérleti csoportban az Ra szignifikánsan meghaladja a kontrollt. Megjegyzendő, hogy a kísérleti csoportokban ez a mutató a kilencedik napra fokozatosan jelentős növekedést mutatott.
A hatások statisztikai megbízhatóságának értékeléséhez az F-kritérium (az átlagos négyzetek aránya) értékeit a két tényező mindegyikére külön-külön (A - a fogva tartás időtartamának faktora; B - a befolyási tényező) és azok kölcsönhatására kapjuk. (A + B), és a kritérium táblázatos értékeit két P=0,05 és P=0,01 szignifikancia szintre hasonlítottuk össze (2. táblázat).
2. táblázat
A hormonális gyógyszerek hatásának varianciaanalízisének eredményei és a fenntartás időtartama a Hydra oligactis ivartalan szaporodásának intenzitására
Fak- Aktuális csoportokban P. táblázat tori Pituitrin Mammophysin Hyfotocin Gonadotropin Prefison Prednizolon 0,05 0,01 A 3,44 1,40 2,27 2,17 3,62 1,30 1,92 2,50 BAN BEN 8,37 4,04 8,09 4,73 8,26 12,70 4,00 7,08 A+B 1,12 0,96 0,56 0,37 1,07 1,03 1,92 2,50 A táblázatból látható, hogy az impakt faktor F ténye 0,05-ös szignifikanciaszint mellett minden kísérleti csoportban nagyobb, mint az F táblázat, 0,01-es szignifikanciaszintnél pedig ilyen kép figyelhető meg a pituitrinnel, hifotocinnal kezelt csoportokban. , prefizon és prednizolon, és a hatás mértéke a prednizolon csoportban a legnagyobb, sokkal nagyobb, mint a pituitrin, a hyfotocin és a prefizon csoportban, amelyek hasonló hatásúak (a tényértékek nagyon közel állnak). Az A és B faktorok kölcsönhatásának hatása nem minden kísérleti csoportban bizonyított.
Az A faktor esetében az Ffact kisebb, mint az Ftable (mindkét szignifikanciaszinten) a mammophysinnel és prednizolonnal kezelt csoportokban. A hyfotocinnal és gonadotropinnal kezelt csoportokban a Fact nagyobb, mint az Ftable P = 0,05-nél, vagyis ennek a faktornak a hatása nem tekinthető meggyőzően bizonyítottnak, ellentétben a pituitrinnel és a prefizonnal végzett kísérleti csoportokkal, ahol az Ffact nagyobb, mint az Ftable. P=0,01-nél és P=0,05-nél.
Minden hormonális gyógyszerek, a gonadotropin mellett különböző mértékben késlelteti az ivartalan szaporodás kezdetét. Ez azonban csak a prefizonnal kezelt csoportban bizonyult statisztikailag szignifikánsnak (P = 0,01). A kísérletben használt hormonális gyógyszerek nem befolyásolják megbízhatóan a fejlődés időtartamát egyetlen vese, az első és a második vese kölcsönös befolyásának megváltoztatása: pituitrin, mammophysin, prefisone, gonadotropin - csak a fejlődő vese kialakult fejrészének jelenlétében; pituitrin, gonadotropin és prednizolon - a fejlődő vesék legalább egy kialakult talpi szakaszának jelenlétében.
Így a hidrák érzékenysége a széleskörű gerincesek hormonvegyületei, és feltételezik, hogy az exogén hormonvegyületek (mint szinergista vagy antagonista) részt vesznek a hidrában rejlő endokrin szabályozó ciklusban.
Bibliográfia
1. Pertseva M.N. A hormonkompetencia fejlesztésének intermolekuláris alapja. L.: Nauka, 1989.
2. Boguta K.K. Néhány morfológiai elvek alacsonyan szervezett idegrendszerek kialakulása az onto- és filogenezisben // Előrelépések modern biológia. M.: Nauka, 1986. T. 101. sz. 3.
3. Ivanova-Kazas A.A. Az állatok ivartalan szaporodása. L., 1971.
4. Nasledov G.A. Az elemi funkcionális feladatok többváltozós megvalósítása és a molekuláris kölcsönhatások rendszerének egyszerűsítése, mint a funkcionális evolúció mintája // Journal of evolutionary biochemistry and physiology. 1991. T. 27. 5. sz.
5. Natochin Yu.V., Breunlich H. Toxikológiai módszerek alkalmazása a vesefunkciók evolúciójának problémájának tanulmányozásában // Journal of evolutionary biochemistry and physiology. 1991. T. 27. 5. sz.
6. Nyikitina S.M. Szteroid hubbub gerinctelen állatokban: Monográfia. L.: Leningrádi Állami Egyetemi Kiadó, 1987.
7. Afonkin S.Yu. Intercelluláris önfelismerés protozoonokban // Results of Science and Technology. M., 1991. T. 9.
8. Prosser L. Az állatok összehasonlító élettana. M.: Mir, 1977. T. 3.
9. Reznikov K.Yu., Nazarevskaya G.D. Az idegrendszer fejlesztési stratégiája onto- és filogenezisben. Hidra // A modern biológia fejlődése. M.: Nauka, 1988. T. 106. 2. szám (5).
10. Sheiman I.M., Balobanova E.F., Gerinctelenek peptidhormonjai // Előrelépések a modern biológiában. M.: Nauka, 1986. T. 101. sz. 2.
11. Etingof R.N. A neuroreceptorok molekuláris szerkezetének vizsgálata. Módszertani megközelítések, evolúciós szempontok // Journal of evolutionary biochemistry and physiology. 1991. T. 27. 5. sz.
12. Highnam K.C., Hill L. A gerinctelenek összehasonlító endokrinológiája // Edward Arnold, 1.





