A baktériumok olyan mikroorganizmusok, amelyek csak egy sejtből állnak. A baktériumok jellegzetes vonása az egyértelműen meghatározott sejtmag hiánya. Ezért nevezik őket „prokariótáknak”, ami azt jelenti, hogy nukleáris mentesek.
Jelenleg a tudomány körülbelül tízezer baktériumfajt ismer, de van egy olyan feltételezés, hogy több mint egymillió baktériumfaj él a Földön. Úgy tartják, hogy a baktériumok a legősibb élőlények a Földön. Szinte mindenhol élnek - vízben, talajban, légkörben és más szervezetek belsejében.
Kinézet
A baktériumok nagyon kicsik, és csak mikroszkóppal láthatók. A baktériumok alakja meglehetősen változatos. A leggyakoribb formák pálcikák, golyók és spirálok.

A rúd alakú baktériumokat "bacilusoknak" nevezik.
A golyók formájában lévő baktériumok coccusok.
A spirál alakú baktériumok a spirilla.
A baktérium alakja határozza meg mobilitását és egy adott felülethez való kötődési képességét.
A baktériumok szerkezete
A baktériumok szerkezete meglehetősen egyszerű. Ezeknek az organizmusoknak több fő szerkezetük van - nukleoid, citoplazma, membrán és sejtfal, emellett sok baktériumnak van flagellája a felszínen.
Nukleoid- Ez olyan, mint egy sejtmag, amely a baktérium genetikai anyagát tartalmazza. Csak egy kromoszómából áll, amely úgy néz ki, mint egy gyűrű.
Citoplazma körülveszi a nukleoidot. A citoplazma fontos struktúrákat tartalmaz - riboszómákat, amelyek szükségesek a baktériumok fehérjeszintetizálásához.
Membrán, kívülről lefedve a citoplazmát, fontos szerepet játszik a baktérium életében. Elhatárolja a baktérium belső tartalmát a külső környezettől, és biztosítja a sejt és a környezet közötti cserefolyamatokat.
A membrán külseje körül van véve sejtfal.
A flagellák száma változhat. Fajtól függően egy baktérium egytől ezer flagellával rendelkezik, de vannak baktériumok nélkül is. A baktériumoknak flagellára van szükségük ahhoz, hogy az űrben mozogjanak.

A baktériumok táplálkozása
A baktériumoknak kétféle táplálkozásuk van. A baktériumok egyik része autotróf, a másik heterotróf.
Az autotrófok kémiai reakciók révén hozzák létre saját tápanyagaikat, míg a heterotrófok más szervezetek által létrehozott szerves anyagokkal táplálkoznak.
Baktériumok szaporodása
A baktériumok osztódással szaporodnak. Az osztódási folyamat előtt a baktérium belsejében található kromoszóma megkétszereződik. Ezután a sejt két részre osztódik. Az eredmény két egyforma leánysejt, amelyek mindegyike megkapja az anya kromoszómájának másolatát.
A baktériumok jelentősége
A baktériumok létfontosságú szerepet játszanak a természetben lévő anyagok körforgásában – a szerves maradékokat szervetlen anyagokká alakítják. Ha nem lennének baktériumok, akkor az egész földet kidőlt fák, lehullott levelek és elhullott állatok borítják.
A baktériumok kettős szerepet játszanak az emberi életben. Egyes baktériumok nagy előnyökkel járnak, míg mások jelentős károkat okoznak.
Számos baktérium kórokozó, és különféle betegségeket okoz, mint például a diftéria, tífusz, pestis, tuberkulózis, kolera és mások.
Vannak azonban olyan baktériumok, amelyek az emberek számára előnyösek. Így élnek baktériumok az emberi emésztőrendszerben, amelyek hozzájárulnak a normál emésztéshez. A tejsavbaktériumokat pedig régóta használják az emberek tejsavtermékek – sajtok, joghurt, kefir stb. A baktériumok fontos szerepet játszanak a zöldségek erjesztésében és az ecet előállításában is.

Rövid tájékoztató a baktériumokról.
Ultravékony metszetek elektronmikroszkópos vizsgálatánál a citoplazma membrán háromrétegű membrán (2 2,5 nm vastag sötét réteget egy világos köztes réteg választ el). Szerkezetében hasonló az állati sejtek plazmalemmájához, és kettős foszfolipidek rétegből áll, beágyazott felülettel és integrált fehérjékkel, mintha áthatolna a membrán szerkezetén. A túlzott növekedéssel (a sejtfal növekedéséhez képest) a citoplazmatikus membrán invaginátumokat képez - invaginációk összetett csavart membránszerkezetek, úgynevezett mezoszómák formájában. A kevésbé bonyolultan csavart struktúrákat intracitoplazmatikus membránoknak nevezzük.
Citoplazma
A citoplazma oldható fehérjékből, ribonukleinsavakból, zárványokból és számos kis szemcsékből - riboszómákból áll, amelyek a fehérjék szintéziséért (transzlációjáért) felelősek. A bakteriális riboszómák mérete körülbelül 20 nm, ülepedési együtthatója pedig 70S, ellentétben az eukarióta sejtekre jellemző 80S riboszómákkal. A riboszómális RNS-ek (rRNS-ek) a baktériumok konzervált elemei (az evolúció „molekuláris órája”). A 16S rRNS a kis riboszomális alegység, a 23S rRNS pedig a nagy riboszomális alegység része. A 16S rRNS vizsgálata a génszisztematika alapja, lehetővé téve az élőlények rokonsági fokának felmérését.
A citoplazma különféle zárványokat tartalmaz glikogén granulátum, poliszacharidok, béta-hidroxi-vajsav és polifoszfátok (volutin) formájában. Tartalékanyagok a baktériumok táplálkozásához és energiaszükségletéhez. A Volutin affinitást mutat az alapfestékekhez, és könnyen kimutatható speciális festési módszerekkel (például Neisser) metakromatikus granulátum formájában. A diftéria bacillusban a volutin szemcsék jellegzetes elrendeződése intenzíven festődő sejtpólusok formájában mutatkozik meg.
Nukleoid
A nukleoid a baktériumok sejtmagjának megfelelője. A baktériumok központi zónájában található, kétszálú DNS formájában, gyűrűbe zárva, és szorosan össze van csomagolva, mint egy labda. A baktériumok magjában, az eukariótáktól eltérően, nincs nukleáris burok, sejtmag és bázikus fehérjék (hisztonok). A baktériumsejt általában egy kromoszómát tartalmaz, amelyet egy gyűrűbe zárt DNS-molekula képvisel.
A nukleoidon kívül, amelyet egy kromoszóma képvisel, a baktériumsejt extrakromoszómális öröklődési faktorokat tartalmaz - plazmidokat, amelyek a DNS kovalensen zárt gyűrűi.
Kapszula, mikrokapszula, nyálka
A kapszula 0,2 mikronnál vastagabb nyálkahártya szerkezet, amely szorosan kapcsolódik a bakteriális sejtfalhoz, és világosan meghatározott külső határokkal rendelkezik. A kapszula patológiás anyagból származó lenyomatkenetekben látható. A tiszta baktériumtenyészetekben a kapszula ritkábban képződik. Speciális kenetfestési módszerekkel észlelik (például Burri-Gins szerint), amelyek negatív kontrasztot hoznak létre a kapszula anyagai között: a tinta sötét hátteret hoz létre a kapszula körül. A kapszula poliszacharidokból (exopoliszacharidokból), néha polipeptidekből áll, például az anthrax bacillusban D-glutaminsav polimerjeiből áll. A kapszula hidrofil, és megakadályozza a baktériumok fagocitózisát. A kapszula antigén: a kapszula elleni antitestek a megnagyobbodást okozzák (kapszuladuzzadási reakció).
Sok baktérium mikrokapszulát alkot – 0,2 mikronnál kisebb vastagságú nyálkahártyaképződményt, amely csak elektronmikroszkóppal detektálható. Meg kell különböztetni a kapszulától a mukoid exopoliszacharidokat, amelyeknek nincs egyértelmű határa. A nyálka vízben oldódik.
A bakteriális exopoliszacharidok részt vesznek az adhézióban (a szubsztrátumokhoz tapadnak). A szintézis mellett
A baktériumok által okozott exopoliszacharidok kialakulásának egy másik mechanizmusa is van: a baktériumok extracelluláris enzimjei a diszacharidokon keresztül. Ennek eredményeként dextránok és levánok képződnek.
Flagella
A bakteriális flagella meghatározza a baktériumsejt mozgékonyságát. A zászlók vékony filamentumok, amelyek a citoplazmatikus membránból származnak, és hosszabbak, mint maga a sejt. A flagella vastagsága 12-20 nm, hossza 3-15 µm. 3 részből állnak: egy spirálszálból, egy kampóból és egy alaptestből, amely speciális korongokkal ellátott rudat tartalmaz (1 pár korong a gram-pozitív baktériumokban és 2 pár korong a gram-negatív baktériumokban). A zászlók a citoplazma membránjához és a sejtfalhoz korongokkal kapcsolódnak. Ez egy villanymotor hatását hozza létre egy motorrúddal, amely forgatja a flagellumot. A zászlók fehérjéből állnak - flagellin (a flagellum - flagellumból); egy H antigén. A flagellin alegységek spirál alakúak.
A különböző fajokhoz tartozó baktériumokban a flagellák száma egytől (monotrich) a Vibrio cholerae-ben a baktérium kerülete mentén több tíz és száz flagelláig terjed (peritrich) Escherichia coliban, Proteusban stb. a sejt vége. Az Amphitriciának egy flagellum vagy egy köteg flagella van a sejt ellentétes végein.
Ittak
A pili (fimbriae, villi) fonalszerű képződmény, vékonyabb és rövidebb (3-10 nm x 0,3-10 µm), mint a flagellák. A Pili a sejtfelszínről nyúlik ki, és a pilin fehérjéből áll, amely antigén aktivitással rendelkezik. Vannak pilusok, amelyek az adhézióért felelősek, vagyis a baktériumok az érintett sejthez való kötődéséért, valamint a táplálkozásért, a víz-só anyagcseréért és a szexuális (F-pili) vagy konjugációs piluszok felelősek. Pili számos – sejtenként több száz. Sejtenként azonban általában 1-3 nemi pilus található: ezeket az úgynevezett „hím” donorsejtek alkotják, amelyek transzmissibilis plazmidokat (F-, R-, Col-plazmidokat) tartalmaznak. A nemi pilusok megkülönböztető jellemzője a speciális „férfi” gömb alakú bakteriofágokkal való kölcsönhatás, amelyek intenzíven adszorbeálódnak a nemi pilusokon.
Vita
A spórák a nyugvó firmicute baktériumok sajátos formája, azaz. baktériumok
Gram-pozitív típusú sejtfalszerkezettel. A spórák a baktériumok létezésére kedvezőtlen körülmények között (száradás, tápanyaghiány stb.) keletkeznek. A baktériumsejt belsejében egy spóra (endospóra) képződik A spórák képződése hozzájárul a faj megőrzéséhez, nem szaporodási módszer A gombákhoz hasonlóan a Bacillus nemzetséghez tartozó spórák nem haladják meg a sejt átmérőjét Azokat a baktériumokat, amelyekben a spóra mérete meghaladja a sejt átmérőjét, pl. lat Clostridium - orsó kék színben.
A spórák alakja lehet ovális, gömb alakú; hely a cellában terminális, azaz. a pálcika végén (a tetanusz kórokozójában), subterminálisan - közelebb a pálcika végéhez (a botulinum kórokozóiban, a gáz gangrénában) és a központi (az anthrax bacillusban). A spóra hosszú ideig fennmarad a többrétegű héj, a kalcium-dipikolinát, az alacsony víztartalom és a lassú anyagcsere-folyamatok miatt. Kedvező körülmények között a spórák kicsíráznak, három egymást követő szakaszon mennek keresztül: aktiválás, iniciáció, csírázás.
A baktériumok morfológiája, a prokarióta sejt szerkezete.
A prokarióta sejtekben nincs egyértelmű határ a sejtmag és a citoplazma között, és nincs magmembrán sem. Ezekben a sejtekben a DNS nem képez az eukarióta kromoszómákhoz hasonló szerkezeteket. Ezért a mitózis és a meiózis folyamatai nem fordulnak elő prokariótákban. A legtöbb prokarióta nem alkot membránokkal határolt intracelluláris organellumokat. Ezenkívül a prokarióta sejtekben nincs mitokondrium vagy kloroplaszt.
Baktériumok, általában egysejtű szervezetek, sejtjük meglehetősen egyszerű alakú, golyó vagy henger, néha ívelt. A baktériumok elsősorban két egyenlő sejtre osztva szaporodnak.
gömb alakú baktériumok hívják cocciés lehet gömb alakú, ellipszoid, bab alakú és lándzsa alakú.
A sejtek osztódás utáni egymáshoz viszonyított elhelyezkedése alapján a coccusok több formára oszlanak. Ha az osztódás után a sejtek eltérnek egymástól, és egyenként helyezkednek el, akkor az ilyen formákat nevezzük monococcusok. Néha a coccusok osztódáskor szőlőfürtre emlékeztető fürtöket képeznek. Hasonló formák utalnak staphylococcus. Azokat a coccusokat, amelyek egy síkban való osztás után összefüggő párokban maradnak, hívjuk diplococcusok, és a különböző lánchosszúságú generátorok azok streptococcusok. Négy coccus kombinációja, amelyek sejtosztódás után jelennek meg két egymásra merőleges síkban tetracoccusok. Egyes coccusok három, egymásra merőleges síkra osztódnak, ami sajátos, köbös alakú, szardíniának nevezett klaszterek kialakulásához vezet.
A legtöbb baktériumnak van hengeres, vagy rúd alakú, alakú. A spórákat alkotó rúd alakú baktériumokat ún bacilusokés nem képeznek spórákat - baktériumok.
A rúd alakú baktériumok alakja, mérete, hosszában és átmérőjében, a sejtvégek alakjában és egymáshoz viszonyított helyzetében is különbözik. Lehetnek henger alakúak, egyenes végekkel vagy oválisak, lekerekített vagy hegyes végekkel. A baktériumok enyhén görbültek is lehetnek, fonalas és elágazó formák találhatók (például mikobaktériumok és aktinomicéták).
Az egyes sejtek osztódás utáni relatív elrendeződésétől függően a pálcika alakú baktériumokat magukra pálcikákra osztják (a sejtek egyetlen elrendezése), diplobaktériumokra vagy diplobacillusokra (sejtek páros elrendezése), streptobakteriákra vagy streptobacillusokra (változó hosszúságú láncokat alkotnak). Gyakran előfordulnak ráncos vagy spirál alakú baktériumok. Ebbe a csoportba tartoznak a spirilla (a latin spira - göndör szóból), amelyek hosszú, ívelt (4-6 fordulatú) rudak, és a vibriók (latinul vibrio - hajlítom), amelyek csak a spirál fordulatának 1/4-ét teszik ki. , hasonló a vesszőhöz.
A baktériumok fonalas formái ismertek, amelyek víztestekben élnek. A felsoroltakon kívül vannak olyan többsejtű baktériumok, amelyek etikus kinövéseket hordoznak a protoplazmatikus sejt felszínén - prostheca, háromszög- és csillag alakú baktériumok, valamint zárt és nyitott gyűrű alakúak és féreg alakú baktériumok.
A baktériumsejtek nagyon kicsik. Mikrométerben mérik, a finom szerkezeti részleteket pedig nanométerben. A coccusok átmérője általában körülbelül 0,5-1,5 mikron. A baktériumok pálca alakú (hengeres) formáinak szélessége a legtöbb esetben 0,5-1 mikron, hossza pedig több mikrométer (2-10). A kis rudak szélessége 0,2-0,4, hossza 0,7-1,5 mikron. A baktériumok között valódi óriások is lehetnek, amelyek hossza eléri a tíz, sőt több száz mikrométert. A baktériumok alakja és mérete jelentősen eltér a tenyészet korától, a táptalaj összetételétől és ozmotikus tulajdonságaitól, hőmérsékletétől és egyéb tényezőktől függően.
A baktériumok három fő formája közül a coccusok a legstabilabbak, a pálcika alakú baktériumok változékonyabbak, a sejthossz különösen jelentősen változik.
A szilárd táptalaj felületére helyezett baktériumsejt növekszik és osztódik, utódbaktériumok kolóniáját képezve. Néhány óra növekedés után a telep már akkora számú sejtből áll, hogy szabad szemmel is látható. A telepek nyálkás vagy pépes állagúak lehetnek, és bizonyos esetekben pigmentáltak. Néha a telepek megjelenése annyira jellemző, hogy lehetővé teszi a mikroorganizmusok különösebb nehézség nélküli azonosítását.
A bakteriális fiziológia alapjai.
Kémiai összetételüket tekintve a mikroorganizmusok alig különböznek más élő sejtektől.
A víz 75-85%-át teszi ki, vegyszerek oldódnak benne.
Szárazanyag 15-25%, szerves és ásványi vegyületeket tartalmaz
A baktériumok táplálkozása. A tápanyagok többféle módon jutnak be a baktériumsejtbe, és függenek az anyagok koncentrációjától, a molekulák méretétől, a környezet pH-jától, a membrán permeabilitásától stb. Ételtípus szerint A mikroorganizmusok a következőkre oszlanak:
autotrófok - szintetizálják az összes széntartalmú anyagot CO2-ból;
heterotrófok – szerves anyagokat használnak szénforrásként;
szaprofiták - elhalt szervezetekből származó szerves anyagokkal táplálkoznak;
A baktériumok légzése. A légzés vagy a biológiai oxidáció olyan redox reakciókon alapul, amelyek egy ATP-molekula képződésével lépnek fel. A molekuláris oxigén tekintetében a baktériumok három fő csoportra oszthatók:
kötelező aerobok - csak oxigén jelenlétében nőhetnek;
kötelező anaerobok - oxigén nélküli közegben nőnek, amely mérgező számukra;
fakultatív anaerobok – növekedhetnek oxigénnel vagy anélkül.
A baktériumok növekedése és szaporodása. A legtöbb prokarióta bináris hasadással szaporodik, ritkábban bimbózás és töredezettség útján. A baktériumokat általában nagy szaporodási arány jellemzi. A különböző baktériumok sejtosztódási ideje meglehetősen széles skálán mozog: E. coli esetében 20 perctől Mycobacterium tuberculosis esetében 14 óráig. A szilárd táptalajokon a baktériumok sejtcsoportokat, úgynevezett kolóniákat alkotnak.
Bakteriális enzimek. Az enzimek fontos szerepet játszanak a mikroorganizmusok anyagcseréjében. Vannak:
endoenzimek - a sejtek citoplazmájában lokalizálódnak;
exoenzimek – kikerülnek a környezetbe.
Az agressziós enzimek elpusztítják a szöveteket és a sejteket, ami a mikrobák és toxinjaik széles körű elterjedését okozza a fertőzött szövetben. A baktériumok biokémiai tulajdonságait az enzimek összetétele határozza meg:
szacharolitikus – a szénhidrátok lebontása;
proteolitikus – fehérjék lebontása,
lipolitikus – zsírbontás,
és fontos diagnosztikai jellemzők a mikroorganizmusok azonosításában.
Számos kórokozó mikroorganizmus esetében az optimális hőmérséklet 37°C és pH 7,2-7,4.
Víz. A víz jelentősége a baktériumok számára. A víz a baktériumok tömegének körülbelül 80%-át teszi ki. A baktériumok növekedése és fejlődése feltétlenül függ a víz jelenlététől, mivel az élő szervezetekben végbemenő összes kémiai reakció vízi környezetben megy végbe. A mikroorganizmusok normális növekedéséhez és fejlődéséhez víz jelenléte szükséges a környezetben.
A baktériumok esetében az aljzat víztartalmának 20%-nál nagyobbnak kell lennie. A víznek hozzáférhető formában kell lennie: folyékony fázisban, 2 és 60 ° C közötti hőmérséklet-tartományban; ezt az intervallumot biokinetikus zónának nevezik. Bár a víz kémiailag nagyon stabil, ionizációs termékei - H+ és OH" ionok igen nagy hatással vannak a sejt szinte minden komponensének (fehérjék, nukleinsavak, lipidek, stb.) tulajdonságaira. Így a katalitikus aktivitás Az enzimek mennyisége nagymértékben függ a H+ és OH ionok koncentrációjától."
A baktériumok energiához jutásának fő módja a fermentáció.
A fermentáció egy anyagcsere-folyamat, amely ATP képződését eredményezi, az elektrondonorok és -akceptorok pedig a fermentáció során keletkező termékek.
A fermentáció szerves anyagok, főként szénhidrátok enzimatikus lebontásának folyamata, amely oxigén felhasználása nélkül megy végbe. Energiaforrásként szolgál a test életéhez, és nagy szerepet játszik az anyagok körforgásában és a természetben. A mikroorganizmusok által kiváltott fermentáció egyes típusait (alkohol, tejsav, vajsav, ecetsav) etil-alkohol, glicerin és egyéb műszaki és élelmiszeripari termékek előállítása során alkalmazzák.
Alkoholos erjesztés(élesztő és bizonyos típusú baktériumok hajtják végre), melynek során a piruvát etanollá és szén-dioxiddá bomlik. Egy glükózmolekula két molekula alkoholt (etanolt) és két molekula szén-dioxidot eredményez. Ez az erjesztési mód nagyon fontos a kenyérgyártásban, a sörfőzésben, a borkészítésben és a lepárlásban.
Tejsavas fermentáció, melynek során a piruvát tejsavvá redukálódik, tejsavbaktériumok és más szervezetek végzik. A tej erjesztésekor a tejsavbaktériumok a laktózt tejsavvá alakítják, így a tejet erjesztett tejtermékekké alakítják (joghurt, aludttej stb.); A tejsav savanyú ízt ad ezeknek a termékeknek.
A tejsavas erjedés az állatok izomzatában is megtörténik, amikor az energiaszükséglet nagyobb, mint a légzés által biztosítotté, és a vérnek nincs ideje oxigént szállítani.
A megerőltető edzés során az izmokban fellépő égő érzés korrelál a tejsavtermeléssel és az anaerob glikolízisre való átállással, mivel az oxigén az aerob glikolízis során gyorsabban alakul szén-dioxiddá, mint ahogy a szervezet oxigént pótol; az edzés utáni izomfájdalmat pedig az izomrostok mikrotrauma okozza. A szervezet oxigénhiány esetén átvált erre a kevésbé hatékony, de gyorsabb ATP-termelési módszerre. A máj ezután megszabadul a felesleges laktáttól, és visszaalakítja azt a fontos glikolitikus intermedier piruváttá.
Ecetsavas fermentáció sok baktérium hajtja végre. Az ecet (ecetsav) a bakteriális fermentáció közvetlen eredménye. Az élelmiszerek pácolása során az ecetsav megvédi az élelmiszereket a patogén és rothadó baktériumoktól.
Vajsav az erjedés vajsav képződéséhez vezet; kórokozói a Clostridium nemzetség néhány anaerob baktériuma.
A baktériumok szaporodása.
Egyes baktériumok nem rendelkeznek ivaros folyamattal, és csak azonos bináris keresztirányú hasadás vagy bimbózással szaporodnak. Az egysejtű cianobaktériumok egyik csoportja esetében többszörös hasadást (gyors, egymást követő bináris osztódások sorozata, amely 4-1024 új sejt kialakulásához vezet) írtak le. Az evolúcióhoz és a változó környezethez való alkalmazkodáshoz szükséges genotípus plaszticitásának biztosítására más mechanizmusokkal rendelkeznek.
Osztódáskor a legtöbb Gram-pozitív baktérium és fonalas cianobaktérium egy keresztirányú septumot szintetizál a perifériától a középpontig a mezoszómák részvételével. A Gram-negatív baktériumok szűkülettel osztódnak: az osztódás helyén a CPM és a sejtfal fokozatosan növekvő befelé görbülését észlelik. Rügyezéskor az anyasejt egyik pólusán rügy képződik és nő az öregedés jeleit mutatja, és általában nem tud 4-nél több leánysejtet termelni. A rügyezés a baktériumok különböző csoportjaiban fordul elő, és feltehetően többször is keletkezett az evolúció során.
Más baktériumokban a szaporodáson kívül a szexuális folyamat is megfigyelhető, de a legprimitívebb formában. A baktériumok nemi folyamata abban különbözik az eukarióták szexuális folyamatától, hogy a baktériumok nem képeznek ivarsejteket, és nem történik sejtfúzió. A rekombináció mechanizmusa prokariótákban. A szexuális folyamat legfontosabb eseménye, a genetikai anyagcsere azonban ebben az esetben is megtörténik. Ezt nevezik genetikai rekombinációnak. A donor sejtből származó DNS egy része (nagyon ritkán az összes DNS) egy olyan recipiens sejtbe kerül, amelynek DNS-e genetikailag különbözik a donor DNS-étől. Ebben az esetben az átvitt DNS helyettesíti a recipiens DNS-ének egy részét. A DNS-csere folyamata olyan enzimeket foglal magában, amelyek felhasítják és újra összekapcsolják a DNS-szálakat. Ez olyan DNS-t termel, amely mindkét szülősejt génjét tartalmazza. Ezt a DNS-t rekombinánsnak nevezik. Az utódok vagy a rekombinánsok a géneltolódások miatt jelentős eltéréseket mutatnak a tulajdonságokban. A karakterek sokfélesége nagyon fontos az evolúció szempontjából, és ez a szexuális folyamat fő előnye.
3 ismert módszer létezik a rekombinánsok előállítására. Ezek - felfedezésük sorrendjében - transzformáció, konjugáció és transzdukció.
A baktériumok eredete.
A baktériumok az archaeákkal együtt az első élő szervezetek közé tartoztak a Földön, körülbelül 3,9-3,5 milliárd évvel ezelőtt jelentek meg. Az e csoportok közötti evolúciós kapcsolatokat még nem vizsgálták teljesen, legalább három fő hipotézis létezik: N. Pace azt sugallja, hogy a protobaktériumok közös ősei vannak. extrém élőhelyeket sajátított el; végül a harmadik hipotézis szerint az archaeák az első élő szervezetek, amelyekből a baktériumok származnak.
Az eukarióták a baktériumsejtek szimbiogenezisének eredményeként jóval később, körülbelül 1,9-1,3 milliárd évvel ezelőtt keletkeztek. A baktériumok evolúcióját kifejezett fiziológiai és biokémiai torzítás jellemzi: az életformák viszonylagos szegénységével és a primitív szerkezettel szinte minden jelenleg ismert biokémiai folyamatot elsajátítottak. A prokarióta bioszférában már megvolt az összes jelenleg létező anyagátalakítási mód. Az eukarióták, miután behatoltak, működésüknek csak mennyiségi vonatkozásait változtatták meg, az elemek körforgásának számos szakaszában azonban nem, a baktériumok továbbra is monopolhelyzetben vannak.
A legrégebbi baktériumok némelyike cianobaktérium. A 3,5 milliárd évvel ezelőtt keletkezett kőzetekben létfontosságú tevékenységük termékeit találták - a cianobaktériumok létezésének vitathatatlan bizonyítékai 2,2-2,0 milliárd évvel ezelőttre nyúlnak vissza. Nekik köszönhetően kezdett felhalmozódni az oxigén a légkörben, amely 2 milliárd évvel ezelőtt elérte az aerob légzés megkezdéséhez elegendő koncentrációt. Az obligát aerob Metallogeniumra jellemző képződmények ebből az időből származnak.
Az oxigén megjelenése a légkörben (oxigénkatasztrófa) komoly csapást mért az anaerob baktériumokra. Vagy kihalnak, vagy helyileg megőrzött oxigénmentes zónákba költöznek. A baktériumok általános fajdiverzitása ekkor csökken.
Feltételezik, hogy a nemi folyamat hiánya miatt a baktériumok evolúciója teljesen más mechanizmust követ, mint az eukariótáké. Az állandó vízszintes géntranszfer kétértelműségekhez vezet az evolúciós összefüggések képében. Az evolúció rendkívül lassan megy végbe (és az eukarióták megjelenésével talán teljesen leáll), de változó körülmények között a gének gyors újraeloszlása zajlik a sejtek között, állandó közös genetikailag; medence.
A baktériumok rendszertana.
A baktériumok szerepe a természetben és az emberi életben.
A baktériumok fontos szerepet játszanak a Földön. Aktívan részt vesznek a természetben lévő anyagok körforgásában. Minden szerves vegyület és a szervetlenek jelentős része jelentős változáson megy keresztül a baktériumok segítségével. Ez a természetben betöltött szerep globális jelentőségű. Miután minden szervezetnél korábban megjelentek a Földön (több mint 3,5 milliárd évvel ezelőtt), létrehozták a Föld élő héját, és továbbra is aktívan dolgozzák fel az élő és elhalt szerves anyagokat, bevonva anyagcseréjük termékeit az anyagok körforgásába. Az anyagok körforgása a természetben az élet létezésének alapja a Földön.
Az összes növényi és állati maradvány lebontását, a humusz és a humusz képződését szintén főként baktériumok végzik. A baktériumok erős biotikus tényező a természetben.
A baktériumok talajképző munkája nagy jelentőséggel bír. Bolygónk első talaját baktériumok hozták létre. A talaj állapota és minősége azonban korunkban is a talajbaktériumok működésétől függ. A talaj termékenysége szempontjából kiemelten fontosak az úgynevezett nitrogénmegkötő göbbaktériumok, a hüvelyes növények szimbiontái. Értékes nitrogénvegyületekkel telítik a talajt.
A baktériumok megtisztítják a szennyvizet a szerves anyagok lebontásával és ártalmatlan szervetlen anyagokká alakításával. A baktériumoknak ezt a tulajdonságát széles körben használják szennyvíztisztító telepeken.
Sok esetben a baktériumok károsak lehetnek az emberre. Így a szaprotróf baktériumok elrontják az élelmiszereket. A termékek romlás elleni védelme érdekében speciális feldolgozásnak vetik alá (forralás, sterilizálás, fagyasztás, szárítás, vegyszeres tisztítás stb.). Ha ezt nem teszik meg, ételmérgezés léphet fel.
A baktériumok között sok olyan betegséget okozó (kórokozó) faj található, amelyek emberekben, állatokban vagy növényekben betegségeket okoznak. A hastífuszt a Salmonella baktérium, míg a vérhasat a Shigella baktérium okozza. A kórokozó baktériumok a levegőn keresztül nyálcseppekkel terjednek a beteg embertől tüsszögéskor, köhögéskor, sőt normál beszélgetés közben is (diftéria, szamárköhögés). Egyes kórokozó baktériumok nagyon ellenállnak a kiszáradásnak, és hosszú ideig megmaradnak porban (tuberculosis bacillus). A Clostridium nemzetséghez tartozó baktériumok porban és talajban élnek - a gáz gangréna és a tetanusz kórokozói. Egyes bakteriális betegségek a beteg személlyel való fizikai érintkezés útján terjednek (szexuális úton terjedő betegségek, lepra). A patogén baktériumokat gyakran úgynevezett vektorok segítségével továbbítják az emberre. Például a legyek a szennyvízben mászva kórokozó baktériumok ezreit gyűjtik össze a lábukon, majd az ember által fogyasztott élelmiszereken hagyják.
Baktériumok a növényi élet egy nagyon egyszerű formája, amely egyetlen élő sejtből áll. A szaporodás sejtosztódással történik. Az érettség elérésekor a baktérium két részre oszlik egyenlő sejtek. Viszont ezek a sejtek mindegyike eléri az érettséget, és szintén két egyenlő sejtre osztódik. Ideális körülmények között baktérium 20-30 percen belül éri el az érettséget és szaporodik. Ilyen szaporodási sebesség mellett egy baktérium elméletileg 34 billió utódot hozhat létre 24 óra alatt! Szerencsére a baktériumok életciklusa viszonylag rövid, néhány perctől néhány óráig tart. Ezért még ideális körülmények között sem tudnak ilyen ütemben szaporodni.Növekedési ütem és bakteriális növekedésés más mikroorganizmusok a környezeti feltételektől függ. A hőmérséklet, a fény, az oxigén, a páratartalom és a pH (savasság vagy lúgosság szintje), valamint a táplálkozás befolyásolja a baktériumok szaporodásának sebességét. Ezek közül a hőmérséklet különösen érdekes a technikusok és mérnökök számára. Minden baktériumtípushoz meg van adva egy minimális hőmérséklet, amelyen szaporodhatnak. E küszöb alatt a baktériumok hibernálnak, és nem képesek szaporodni. Mindegyiknél pontosan ugyanaz baktériumok típusai van egy maximális hőmérsékleti küszöb. E határ feletti hőmérsékleten a baktériumok elpusztulnak. E határok között van az az optimális hőmérséklet, amelyen a baktériumok maximális sebességgel szaporodnak. A legtöbb baktérium számára, amelyek állati ürülékkel, valamint elhalt állati és növényi szövetekkel (szaprofitákkal) táplálkoznak, az optimális hőmérséklet 24-30°C. A legtöbb baktérium számára, amelyek fertőzéseket és betegségeket okoznak a gazdaszervezetben (kórokozó baktériumok), körülbelül 38 °C. A legtöbb esetben jelentősen csökkentheti bakteriális növekedési sebesség, ha a környezet. Végül, számos baktériumfajta létezik, amelyek vízhőmérsékleten, míg mások fagyos hőmérsékleten fejlődnek a legjobban.
Kiegészítés a fentiekhez
Eredet, evolúció, hely a földi élet kialakulásában
A baktériumok az archaeákkal együtt az első élő szervezetek közé tartoztak a Földön, körülbelül 3,9-3,5 milliárd évvel ezelőtt jelentek meg. Az e csoportok közötti evolúciós kapcsolatokat még nem vizsgálták teljesen, legalább három fő hipotézis létezik: N. Pace azt sugallja, hogy a protobaktériumok közös ősei vannak. extrém élőhelyeket sajátított el; végül a harmadik hipotézis szerint az archaeák az első élő szervezetek, amelyekből a baktériumok származnak.Az eukarióták a baktériumsejtek szimbiogenezisének eredményeként jóval később, körülbelül 1,9-1,3 milliárd évvel ezelőtt keletkeztek. A baktériumok evolúcióját kifejezett fiziológiai és biokémiai torzítás jellemzi: az életformák viszonylagos szegénységével és a primitív szerkezettel szinte minden jelenleg ismert biokémiai folyamatot elsajátítottak. A prokarióta bioszférában már megvolt az összes jelenleg létező anyagátalakítási mód. Az eukarióták, miután behatoltak, működésüknek csak mennyiségi vonatkozásait változtatták meg, az elemek minőségi szempontjait azonban nem, a baktériumok továbbra is monopolhelyzetben vannak.
A legrégebbi baktériumok némelyike cianobaktérium. A 3,5 milliárd évvel ezelőtt keletkezett kőzetekben létfontosságú tevékenységük termékeit találták - a cianobaktériumok létezésének vitathatatlan bizonyítékai 2,2-2,0 milliárd évvel ezelőttre nyúlnak vissza. Nekik köszönhetően kezdett felhalmozódni az oxigén a légkörben, amely 2 milliárd évvel ezelőtt elérte az aerob légzés megkezdéséhez elegendő koncentrációt. Az obligát aerob Metallogeniumra jellemző képződmények ebből az időből származnak.
Az oxigén megjelenése a légkörben komoly csapást mért az anaerob baktériumokra. Vagy kihalnak, vagy helyileg megőrzött oxigénmentes zónákba költöznek. A baktériumok általános fajdiverzitása ekkor csökken.
Feltételezik, hogy a nemi folyamat hiánya miatt a baktériumok evolúciója teljesen más mechanizmust követ, mint az eukariótáké. Az állandó vízszintes géntranszfer kétértelműségekhez vezet az evolúciós összefüggések képében. Az evolúció rendkívül lassan megy végbe (és az eukarióták megjelenésével talán teljesen leáll), de változó körülmények között a gének gyors újraeloszlása zajlik a sejtek között, állandó közös genetikailag; medence.
Szerkezet
A baktériumok túlnyomó többsége (az aktinomyceták és a fonalas cianobaktériumok kivételével) egysejtű. A sejtek alakja szerint lehetnek kerekek (coccusok), rúd alakúak (bacilusok, klostrídiumok, pszeudomonádok), csavartak (vibriók, spirillumok, spirocheták), ritkábban csillagok, tetraéderek, köbösek, C- vagy O- alakú. Az alak határozza meg a baktériumok képességeit, például a felülethez való kötődést, a mobilitást és a tápanyagok felszívódását. Megfigyelték például, hogy az oligotrófok, vagyis a környezetben alacsony tápanyagtartalmú baktériumok a felület-térfogat arány növelésére törekszenek például kinövések (ún. prostek) képződésével. ).A kötelező sejtszerkezetek közül hármat különböztetünk meg:
- nukleoid
- riboszómák
- citoplazmatikus membrán (CPM)
Protoplaszt szerkezet
A CPM korlátozza a sejt (citoplazma) tartalmát a külső környezetből. A citoplazma homogén frakcióját, amely oldható RNS-t, fehérjéket, metabolikus reakciók termékeit és szubsztrátjait tartalmazza, citoszolnak nevezzük. A citoplazma másik részét különféle szerkezeti elemek képviselik.Az egyik fő különbség a baktériumsejt és az eukarióta sejt között a nukleáris membrán hiánya, és szigorúan véve az intracitoplazmatikus membránok általános hiánya, amelyek nem a CPM származékai. A prokarióták különböző csoportjaiban (különösen gyakran a gram-pozitív baktériumokban) azonban a CPM-mezoszómák helyi invaginációi vannak, amelyek különféle funkciókat látnak el a sejtben, és funkcionálisan különböző részekre osztják fel. Számos fotoszintetikus baktérium rendelkezik a CPM-ből származó fotoszintetikus membránok fejlett hálózatával. A lila baktériumokban megőrizték a kapcsolatot a CPM-mel, ami elektronmikroszkóp alatt metszetekben könnyen kimutatható, cianobaktériumokban ez a kapcsolat vagy nehezen észlelhető, vagy az evolúció során elveszett. A tenyésztés körülményeitől és korától függően a fotoszintetikus membránok különféle struktúrákat - hólyagokat, kromatoforokat, tilakoidokat - alkotnak.
A baktériumok életéhez szükséges összes genetikai információ egy DNS-ben (bakteriális kromoszómában) található, leggyakrabban kovalensen zárt gyűrű formájában (a lineáris kromoszómák a Streptomyces és a Borrelia törzsekben találhatók). Egy ponton a CPM-hez kapcsolódik, és egy különálló, de a citoplazmától membránnal el nem választott szerkezetben helyezkedik el, és nukleoidnak nevezik. A kibontott DNS több mint 1 mm hosszú. A bakteriális kromoszóma általában egyetlen példányban jelenik meg, vagyis szinte minden prokarióta haploid, bár bizonyos körülmények között egy sejt több kópiát is tartalmazhat kromoszómájából, a Burkholderia cepacia pedig három különböző gyűrűkromoszómával rendelkezik (3,6, 3,2 és 1,1 millió hosszúságú). nukleotidpárok). A prokarióták riboszómái szintén különböznek az eukariótákétól, és ülepedési állandójuk 70 S (eukariótákban 80 S).
Ezeken a struktúrákon kívül tartalék anyagok zárványai is jelen lehetnek a citoplazmában.
Sejtmembrán és felületi struktúrák
A sejtfal a baktériumsejt fontos szerkezeti eleme, de nem nélkülözhetetlen. Mesterségesen előállították a részben vagy teljesen hiányzó sejtfalú formákat (L-formák), amelyek kedvező körülmények között létezhettek, de néha elvesztették osztódási képességüket. A természetes baktériumok egy olyan csoportja is ismert, amelyek nem tartalmaznak sejtfalat - a mikoplazmák.A baktériumokban a sejtfal szerkezetének két fő típusa van, amelyek a gram-pozitív és gram-negatív fajokra jellemzőek.
A Gram-pozitív baktériumok sejtfala egy 20-80 nm vastag homogén réteg, amely főként peptidoglikánból épül fel, kisebb mennyiségű teichoinsavval és kis mennyiségű poliszacharidokkal, fehérjékkel és lipidekkel (ún. lipopoliszacharid). A sejtfalon 1-6 nm átmérőjű pórusok találhatók, amelyek számos molekula számára áteresztővé teszik.
Gram-negatív baktériumokban a peptidoglikán réteg lazán szomszédos a CPM-mel, és csak 2-3 nm vastagságú. Külső membrán veszi körül, amely általában egyenetlen, ívelt alakú. A CPM, a peptidoglikán réteg és a külső membrán között van egy periplazmatikus tér, amelyet transzportfehérjéket és enzimeket tartalmazó oldat tölt ki.
A sejtfal külső oldalán kapszula lehet - egy amorf réteg, amely fenntartja a kapcsolatot a fallal. A nyálkahártya rétegei nem kapcsolódnak a sejthez, könnyen elválaszthatók, a fedők pedig nem amorfok, hanem finom szerkezetűek. E három idealizált eset között azonban számos átmeneti forma létezik.
0-tól 1000-ig terjedő bakteriális flagella lehet. A lehetséges opciók közé tartozik egy flagellum elrendezése egy póluson (monopoláris monotrich), egy köteg flagella egy póluson (monopoláris peritrichus vagy lophotrichiális flagella) vagy két pólus (bipoláris peritrichus vagy amfitrichiás flagella). valamint számos flagella a sejt teljes felületén (peritrich). A flagellum vastagsága 10-20 nm, hossza - 3-15 µm. Forgása az óramutató járásával ellentétes irányban történik, 40-60 fordulat/perc frekvenciával.
A baktériumok felszíni struktúrái közül a flagellák mellett meg kell említeni a bolyhokat. Vékonyabbak, mint a flagellák (átmérője 5-10 nm, hossza legfeljebb 2 µm), és szükségesek a baktériumok szubsztráthoz való rögzítéséhez, részt vesznek az anyagcseretermékekben, és speciális bolyhok - F-pili - fonalszerű képződmények, vékonyabbak és rövidebbek ( 3-10 nm x 0, 3-10 µm), mint a flagella - ezek szükségesek ahhoz, hogy a donor sejt DNS-t vigyen át a recipiensnek a konjugáció során.
Méretek
A baktériumok átlagos mérete 0,5-5 mikron. Az Escherichia coli mérete például 0,3-1 x 1-6 mikron, a Staphylococcus aureus átmérője 0,5-1 mikron, a Bacillus subtilis 0,75 x 2-3 mikron. A legnagyobb ismert baktérium a Thiomargarita namibiensis, mérete eléri a 750 mikron (0,75 mm) méretet. A második az Epulopiscium fishelsoni, amely 80 mikron átmérőjű és legfeljebb 700 mikron hosszúságú, és az Acanthurus nigrofuscus sebészeti hal emésztőrendszerében él. Az Achromatium oxaliferum eléri a 33 x 100 mikron, a Beggiatoa alba a 10 x 50 mikron méretet. A spirocheták akár 250 µm hosszúra is megnőhetnek, vastagságuk pedig 0,7 µm. Ugyanakkor a baktériumok közé tartoznak a legkisebb sejtszerkezetű organizmusok is. A Mycoplasma mycoides mérete 0,1-0,25 mikron, ami hasonló a nagy vírusok méretéhez, mint például a dohánymozaik, a tehénhimlő vagy az influenza. Elméleti számítások szerint egy 0,15-0,20 mikronnál kisebb átmérőjű gömb alakú sejt önálló szaporodásra képtelenné válik, mivel fizikailag nem tartalmazza kellő mennyiségben az összes szükséges biopolimert és szerkezetet.Leírtak azonban olyan nanobaktériumokat, amelyek kisebbek az „elfogadható” méretnél, és nagyon különböznek a közönséges baktériumoktól. A vírusokkal ellentétben képesek önálló növekedésre és szaporodásra (rendkívül lassú). Eddig kevéssé tanulmányozták őket, élő természetüket megkérdőjelezik.
A sejt sugarának lineáris növekedésével a felülete a sugár négyzetével, a térfogata pedig a kockával arányosan növekszik, ezért a kis szervezetekben a felület és a térfogat aránya nagyobb, mint a nagyobbakban, ami az előbbiek számára a környezettel való aktívabb anyagcserét jelenti. A különböző mutatókkal mért anyagcsere-aktivitás biomassza egységenként nagyobb a kis formákban, mint a nagyoknál. Ezért a kis méretek még mikroorganizmusok számára is előnyöket biztosítanak a baktériumoknak és az archaeáknak a növekedési és szaporodási sebességben a bonyolultabb eukariótákhoz képest, és meghatározzák fontos ökológiai szerepüket.
Többsejtűség a baktériumokban
Az egysejtű formák képesek ellátni a szervezetben rejlő összes funkciót, függetlenül a szomszédos sejtektől. Sok egysejtű prokarióta hajlamos sejtes prokarióták kialakítására, amelyeket gyakran az általuk kiválasztott nyálka tartja össze. Leggyakrabban ez csak az egyes organizmusok véletlenszerű társulása, de bizonyos esetekben átmeneti társulás társul egy bizonyos funkció végrehajtásához, például a myxobaktériumok általi termőtestek kialakulása lehetővé teszi ciszták kialakulását, bár egyes esetekben a sejtek nem képesek kialakítani őket. Az ilyen jelenségek, valamint az egysejtű eubaktériumok által morfológiailag és funkcionálisan differenciált sejtek kialakulása szükséges előfeltételei a valódi többsejtűség kialakulásának.A többsejtű szervezetnek meg kell felelnie a következő feltételeknek:
- sejtjeit aggregálni kell,
- meg kell osztani a funkciókat a sejtek között,
- stabil specifikus kapcsolatokat kell kialakítani az aggregált cellák között.
Baktériumok szaporodása
Egyes baktériumok nem rendelkeznek ivaros folyamattal, és csak azonos bináris keresztirányú hasadás vagy bimbózással szaporodnak. Az egysejtű cianobaktériumok egyik csoportja esetében többszörös hasadást (gyors, egymást követő bináris osztódások sorozata, amely 4-1024 új sejt kialakulásához vezet) írtak le. Az evolúcióhoz és a változó környezethez való alkalmazkodáshoz szükséges genotípus plaszticitásának biztosítására más mechanizmusokkal rendelkeznek.Osztódáskor a legtöbb Gram-pozitív baktérium és fonalas cianobaktérium egy keresztirányú septumot szintetizál a perifériától a középpontig a mezoszómák részvételével. A Gram-negatív baktériumok szűkülettel osztódnak: az osztódás helyén a CPM és a sejtfal fokozatosan növekvő befelé görbülését észlelik. Rügyezéskor az anyasejt egyik pólusán rügy képződik és nő az öregedés jeleit mutatja, és általában nem tud 4-nél több leánysejtet termelni. A rügyezés a baktériumok különböző csoportjaiban fordul elő, és feltehetően többször is keletkezett az evolúció során.
A baktériumok ivaros szaporodást is mutatnak, de a legprimitívebb formában. A baktériumok ivaros szaporodása abban különbözik az eukarióták szexuális szaporodásától, hogy a baktériumok nem képeznek ivarsejteket, és nem mennek keresztül sejtfúzión. Az ivaros szaporodás legfontosabb eseménye, a genetikai anyagcsere azonban ebben az esetben is bekövetkezik. Ezt a folyamatot genetikai rekombinációnak nevezik. A donor sejtből származó DNS egy része (nagyon ritkán az összes DNS) egy olyan recipiens sejtbe kerül, amelynek DNS-e genetikailag különbözik a donor DNS-étől. Ebben az esetben az átvitt DNS helyettesíti a recipiens DNS-ének egy részét. A DNS-csere folyamata olyan enzimeket foglal magában, amelyek felhasítják és újra összekapcsolják a DNS-szálakat. Ez olyan DNS-t termel, amely mindkét szülősejt génjét tartalmazza. Ezt a DNS-t rekombinánsnak nevezik. Az utódokban vagy a rekombinánsokban a géneltolódások által okozott tulajdonságok jelentős eltéréseket mutatnak. A tulajdonságok sokfélesége nagyon fontos az evolúció szempontjából, és a szexuális szaporodás fő előnye. 3 ismert módszer létezik a rekombinánsok előállítására. Ezek - felfedezésük sorrendjében - transzformáció, konjugáció és transzdukció.
A baktériumok hasadással történő szaporodása a leggyakoribb módszer a mikrobiális populáció méretének növelésére. Az osztódás után a baktériumok eredeti méretükre nőnek, amihez bizonyos anyagokra (növekedési faktorokra) van szükség.
A baktériumok szaporodási módjai eltérőek, de a legtöbb fajuk hasadás útján ivartalan szaporodást mutat. A baktériumok rendkívül ritkán szaporodnak bimbózás útján. A baktériumok szexuális szaporodása primitív formában van jelen.
Rizs. 1. A fotón egy baktériumsejt látható az osztódás stádiumában.
A baktériumok genetikai berendezése
A baktériumok genetikai berendezését egyetlen DNS - kromoszóma képviseli. A DNS körbe záródik. A kromoszóma egy membránnal nem rendelkező nukleotidban lokalizálódik. A baktériumsejt plazmidokat tartalmaz.
Nukleoid
A nukleoid a mag analógja. A cella közepén található. Összehajtott formában tartalmazza a DNS-t, az örökletes információ hordozóját. A feltekercselt DNS eléri az 1 mm hosszúságot. A baktériumsejt nukleáris anyagának nincs membránja, sejtmagja vagy kromoszómakészlete, és nem osztódik mitózissal. Osztás előtt a nukleotidot megkétszerezzük. Az osztódás során a nukleotidok száma 4-re nő.

Rizs. 2. A fotón egy baktériumsejt látható egy metszetben. A központi részben egy nukleotid látható.
Plazmidok
A plazmidok autonóm molekulák, amelyek kettős szálú DNS-gyűrűvé vannak hajtva. Tömegük lényegesen kisebb, mint egy nukleotid tömege. Annak ellenére, hogy az örökletes információkat a plazmidok DNS-e kódolja, ezek nem létfontosságúak és nem szükségesek a baktériumsejt számára.

Rizs. 3. A fotón egy bakteriális plazmid látható.
A felosztás szakaszai
A felnőtt sejtre jellemző bizonyos méret elérése után az osztódási mechanizmusok elindulnak.
DNS replikáció
A DNS replikációja megelőzi a sejtosztódást. A mezoszómák (a citoplazma membrán redői) tartják a DNS-t, amíg az osztódási (replikációs) folyamat be nem fejeződik.
A DNS-replikáció a DNS-polimeráz enzimek segítségével történik. A replikáció során a kétszálú DNS-ben a hidrogénkötések megszakadnak, aminek eredményeként egy DNS-ből két egyszálú leány-DNS jön létre. Ezt követően, amikor a leány DNS-ek átvették a helyüket az elválasztott leánysejtekben, helyreállnak.
Amint a DNS-replikáció befejeződött, a szintézis eredményeként szűkület jelenik meg, amely a sejtet felére osztja. Először a nukleotid osztódik, majd a citoplazma. A sejtfal szintézise befejezi az osztódást.
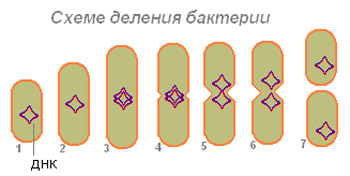
Rizs. 4. A bakteriális sejtosztódás sémája.
DNS szakaszok cseréje
A Bacillus subtilisben a DNS-replikáció folyamata két DNS-szakasz kicserélésével ér véget.
A sejtosztódás után egy híd képződik, amelyen keresztül az egyik sejt DNS-e átjut a másikba. Ezután mindkét DNS összefonódik. A két DNS egyes szakaszai összetapadnak. Az adhézió helyén a DNS-szegmensek kicserélődnek. Az egyik DNS a jumper mentén visszamegy az első sejtbe.

Rizs. 5. A DNS-csere változata Bacillus subtilisben.
A bakteriális sejtosztódás típusai
Ha a sejtosztódás megelőzi az elválasztási folyamatot, akkor többsejtű rudak és coccusok képződnek.
Szinkron sejtosztódással két teljes értékű leánysejt képződik.
Ha a nukleotid gyorsabban osztódik, mint maga a sejt, akkor többnukleotidos baktériumok képződnek.
A baktériumok elválasztásának módszerei
Osztás töréssel
A töréssel való osztódás az antraxbacillusokra jellemző. Ennek az osztódásnak a következtében a sejtek a találkozási pontokon eltörnek, megszakítva a citoplazmahidakat. Aztán taszítják egymást, láncokat alkotva.
Csúszó osztás
Csúszó elválasztásnál az osztódás után a sejt levál, és mintegy végigcsúszik egy másik sejt felületén. Ez az elválasztási módszer az Escherichia egyes formáira jellemző.
Split split
Szekáns osztódás esetén az egyik osztott sejt szabad végével egy körívet ír le, amelynek középpontja egy másik sejttel való érintkezési pontja, és egy római quinque-t vagy ékírást (Corynebacterium diphtheria, Listeria) alkot.

Rizs. 6. A képen rúd alakú baktériumok láthatók, amelyek láncokat (anthraxbacillusokat) képeznek.

Rizs. 7. A képen egy csúszó módszer látható az E. coli elválasztására.

Rizs. 8. A corynebaktériumok szétválasztásának módszere.
A bakteriális klaszterek típusa osztódás után
Az osztódó sejtcsoportok változatos alakúak, amelyek az osztódási sík irányától függenek.
Globuláris baktériumok egyenként, két-kettővel (diplococcusok), csomagokba, láncokba vagy szőlőfürtszerűen rendezve. Rúd alakú baktériumok - láncokban.
Spirál alakú baktériumok- kaotikus.

Rizs. 9. A fotón mikrococcusok láthatók. Kerek, sima, fehér, sárga és piros színűek. A természetben a mikrococcusok mindenütt jelen vannak. Az emberi test különböző üregeiben élnek.

Rizs. 10. A képen diplococcus baktériumok - Streptococcus pneumoniae.

Rizs. 11. A képen Sarcina baktériumok láthatók. A coccoid baktériumok csomagokban csoportosulnak össze.

Rizs. 12. A képen streptococcus baktériumok láthatók (a görög „streptos” - láncból). Láncba rendezve. Számos betegség kórokozói.

Rizs. 13. A fotón a baktériumok „arany” staphylococcusok. Úgy elrendezve, mint „szőlőfürtök”. A fürtök arany színűek. Számos betegség kórokozói.

Rizs. 14. A képen a feltekert Leptospira baktériumok számos betegség kórokozói.

Rizs. 15. A képen a Vibrio nemzetséghez tartozó pálcika alakú baktériumok láthatók.
A baktériumok osztódási sebessége
A baktériumok osztódási sebessége rendkívül magas. Átlagosan 20 percenként osztódik egy baktériumsejt. Egy sejtből egy nap alatt 72 utódnemzedék jön létre. A Mycobacterium tuberculosis lassan osztódik. A teljes felosztási folyamat körülbelül 14 órát vesz igénybe.

Rizs. 16. A fotó a streptococcus sejtosztódás folyamatát mutatja be.
A baktériumok szexuális szaporodása
1946-ban a tudósok felfedezték a nemi szaporodást primitív formában. Ebben az esetben ivarsejtek (hím és női nemi sejtek) nem képződnek, de egyes sejtek genetikai anyagot cserélnek ( genetikai rekombináció).
Ennek eredményeként génátvitel történik konjugáció- a genetikai információ egy részének egyirányú átvitele a formában plazmidok bakteriális sejtekkel való érintkezéskor.
A plazmidok kis DNS-molekulák. Nem kapcsolódnak a kromoszóma genomhoz, és képesek autonóm megkettőződésre. A plazmidok olyan géneket tartalmaznak, amelyek növelik a baktériumsejtek rezisztenciáját a kedvezőtlen környezeti feltételekkel szemben. A baktériumok gyakran adják át egymásnak ezeket a géneket. A genetikai információ átadása más fajba tartozó baktériumok számára is megfigyelhető.
Valódi szexuális folyamat hiányában a konjugáció játszik óriási szerepet a hasznos tulajdonságok cseréjében. Így közvetítik a baktériumok azon képességét, hogy gyógyszerrezisztenciát mutassanak ki. Az antibiotikum-rezisztencia átvitele a betegséget okozó populációk között különösen veszélyes az emberiség számára.

Rizs. 17. A képen két E. coli konjugációjának pillanata látható.
A baktériumpopuláció fejlődésének fázisai
Táptalajra oltva a baktériumpopuláció fejlődése több fázison megy keresztül.
Kezdeti fázis
A kezdeti szakasz a vetés pillanatától a növekedésig tartó időszak. A kezdeti szakasz átlagosan 1-2 óráig tart.
Tenyésztési késleltetési szakasz
Ez az intenzív baktériumnövekedés fázisa. Időtartama körülbelül 2 óra. Ez függ a termés korától, az alkalmazkodási időszaktól, a táptalaj minőségétől stb.
Logaritmikus fázis
Ebben a fázisban a szaporodás csúcssebessége és a baktériumpopuláció növekedése tapasztalható. Időtartama 5-6 óra.
Negatív gyorsulási fázis
Ebben a fázisban csökken a szaporodási sebesség, csökken az osztódó baktériumok száma, és nő az elpusztult baktériumok száma. A negatív gyorsulás oka a tápközeg kimerülése. Időtartama körülbelül 2 óra.
Álló maximális fázis
Az állófázisban egyenlő számú elhullott és újonnan képződött egyed figyelhető meg. Időtartama körülbelül 2 óra.
Halálgyorsítási fázis
Ebben a fázisban az elhalt sejtek száma fokozatosan növekszik. Időtartama körülbelül 3 óra.
Logaritmikus halálfázis
Ebben a fázisban a baktériumsejtek állandó sebességgel pusztulnak el. Időtartama körülbelül 5 óra.
Csökkentési ütem
Ebben a fázisban a fennmaradó élő baktériumsejtek alvó állapotba kerülnek.

Rizs. 18. Az ábra egy baktériumpopuláció növekedési görbéjét mutatja.

Rizs. 19. A képen a Pseudomonas aeruginosa telepe kékeszöld, a micrococcusok telepe sárga, a Bacterium prodigiosum kolónia vérvörös, a Bacteroides niger kolónia pedig fekete.

Rizs. 20. A fotón egy baktériumkolónia látható. Minden kolónia egyetlen sejt utódai. Egy kolóniában a sejtek száma milliós nagyságrendű. A telep 1-3 nap alatt nő.
A mágnesesre érzékeny baktériumok osztódása
Az 1970-es években felfedezték a tengerekben élő baktériumokat, amelyeknek mágneses érzetük volt. A mágnesesség lehetővé teszi ezeknek a csodálatos lényeknek, hogy a Föld mágneses mezejének vonalai mentén eligazodjanak, és találjanak ként, oxigént és más olyan anyagokat, amelyekre annyira szüksége van. Az ő „iránytűjüket” a magnetoszómák képviselik, amelyek mágnesből állnak. Osztásakor a mágnesesre érzékeny baktériumok szétosztják iránytűjüket. Ilyenkor az osztódás közbeni szűkület egyértelműen elégtelenné válik, így a baktériumsejt meghajlik és éles törést okoz.

Rizs. 21. A fotón egy mágnesesen érzékeny baktérium osztódási pillanata látható.
Bakteriális növekedés
Amikor egy baktériumsejt elkezd osztódni, két DNS-molekula a sejt ellentétes végeire költözik. Ezután a cellát két egyenlő részre osztják, amelyeket elválasztanak egymástól, és eredeti méretükre növelik. Sok baktérium osztódási sebessége átlagosan 20-30 perc. Egy sejtből egy nap alatt 72 utódnemzedék jön létre.
A növekedési és fejlődési folyamat során egy sejttömeg gyorsan felszívja a tápanyagokat a környezetből. Ezt elősegítik a kedvező környezeti tényezők - a hőmérsékleti viszonyok, a megfelelő mennyiségű tápanyag, valamint a környezet szükséges pH-értéke. Az aerob sejteknek oxigénre van szükségük. Veszélyes az anaerobokra. A baktériumok korlátlan szaporodása azonban nem fordul elő a természetben. A napfény, a száraz levegő, a táplálékhiány, a magas környezeti hőmérséklet és egyéb tényezők káros hatással vannak a baktériumsejtre.

Rizs. 22. A képen a sejtosztódás pillanata látható.
Növekedési tényezők
A baktériumok szaporodásához bizonyos anyagok (növekedési faktorok) szükségesek, amelyek egy részét maga a sejt szintetizálja, egy részük a környezetből származik. A növekedési faktorok szükségessége minden baktérium esetében eltérő.
A növekedési faktorok iránti igény állandó jellemző, amely lehetővé teszi a baktériumok azonosítására, táptalajok készítésére és biotechnológiai felhasználására.
A bakteriális növekedési faktorok (bakteriális vitaminok) kémiai elemek, amelyek többsége vízben oldódó B-vitamin Ebbe a csoportba tartoznak még a hemin, kolin, purin és pirimidin bázisok és egyéb aminosavak. Növekedési faktorok hiányában bakteriosztázis lép fel.
A baktériumok minimális mennyiségben és változatlan formában használnak növekedési faktorokat. Az ebbe a csoportba tartozó számos vegyi anyag a sejtenzimek részét képezi.

Rizs. 23. A fotón egy pálcika alakú baktérium osztódásának pillanata látható.
A legfontosabb bakteriális növekedési faktorok
- B1-vitamin (tiamin). Részt vesz a szénhidrát anyagcserében.
- B2-vitamin (riboflavin). Részt vesz a redox reakciókban.
- Pantoténsav az A koenzim összetevője.
- B6-vitamin (piridoxin). Részt vesz az aminosav-anyagcserében.
- B12 vitaminok(a kobalaminok kobaltot tartalmazó anyagok). Aktívan részt vesznek a nukleotidok szintézisében.
- Folsav. Egyes származékai olyan enzimek részét képezik, amelyek katalizálják a purin- és pirimidinbázisok, valamint néhány aminosav szintézisét.
- Biotin. Részt vesz a nitrogén anyagcserében és katalizálja a telítetlen zsírsavak szintézisét is.
- PP vitamin(nikotinsav). Részt vesz a redox reakciókban, az enzimek képződésében, valamint a lipidek és szénhidrátok anyagcseréjében.
- H vitamin(para-amino-benzoesav). Számos baktérium növekedési faktora, beleértve az emberi belekben élőket is. A folsavat para-amino-benzoesavból szintetizálják.
- Gemin. Egyes enzimek alkotórésze, amelyek részt vesznek az oxidációs reakciókban.
- Kholin. Részt vesz a sejtfal lipidszintézisének reakcióiban. A metilcsoport szállítója az aminosavak szintézisében.
- Purin és pirimidin bázisok(adenin, guanin, xantin, hipoxantin, citozin, timin és uracil). Az anyagokra főleg nukleinsavak komponenseiként van szükség.
- Aminosavak. Ezek az anyagok a sejtfehérjék összetevői.
Egyes baktériumok növekedési faktorainak szükséglete
Auxotrófok Az élet biztosításához szükséges a vegyszerek kívülről történő ellátása. Például a klostrídiumok nem képesek lecitin és tirozin szintetizálására. A staphylococcusok lecitint és arginint igényelnek. A streptococcusok zsírsavakat igényelnek - a foszfolipidek összetevőit. A Corynebacteriumok és a Shigella baktériumok nikotinsavat igényelnek. A Staphylococcus aureus, a pneumococcusok és a Brucella B1-vitamint igényel. Streptococcusok és tetanuszbacilusok - pantoténsavban.
Prototrófokönállóan szintetizálja a szükséges anyagokat.

Rizs. 24. A különböző környezeti feltételek eltérő hatással vannak a baktériumkolóniák növekedésére. A bal oldalon egyenletes növekedés látható, lassan táguló kör formájában. A jobb oldalon a gyors növekedés „hajtások” formájában.
A baktériumok növekedési faktorok iránti igényének tanulmányozása lehetővé teszi a tudósok számára, hogy nagy mikrobiális tömeget érjenek el, amely annyira szükséges az antimikrobiális gyógyszerek, szérumok és vakcinák előállításához.
Tudjon meg többet a baktériumokról a cikkekben:
A baktériumok szaporodása a mikrobapopulációk számának növelésének mechanizmusa. A bakteriális osztódás a szaporodás fő módja. Az osztódás után a baktériumoknak el kell érniük a felnőtt méretet. A baktériumok úgy szaporodnak, hogy gyorsan felszívják a tápanyagokat a környezetükből. A növekedéshez bizonyos anyagokra (növekedési faktorokra) van szükség, amelyek egy részét maga a baktériumsejt szintetizálja, más része pedig a környezetből származik.





