Гидры - это особый род сидячих кишечнополостных, которые своим внешним видом и образом жизни напоминают растения, но все же они относятся к царству животных. Нервная система у гидры устроена таким образом, чтобы обеспечить возможность существу добывать достаточное количество пищи.
Разобраться какого типа нервная система у гидры непросто, так как эта структура довольно проста и встречается не только у этих существ, но также у некоторых видов медуз и других примитивных животных. Гидры - это сравнительно небольшие животные организмы, достигающие размеров от 2 до 20 мм.
Клетки, формирующие нервную систему, по форме напоминают звездочки, которые соединены лучами между собой, образовывая нейронную сеть. Нервная система располагается под кожно-мускульными клетками. Органа центрального восприятия электрических импульсов, вызванных внешними или внутренними раздражителями, у гидр нет. Максимальное количество нейронов составляет примерно 5000 шт. и все они соединены между собой.
Нервная система гидры получила название диффузного плексуса, так как имеет место рассеянное и неоднородное сплетение. Сгущение диффузного плексуса наблюдается в области подошвы, ротовой полости и щупалец. Последние исследования показали, что в области ротового отверстия имеется нервное кольцо, которое отличается схожей структурой с нервным кольцом, располагающимся по краю зонтика гидромедузы.
Нервная система гидры крайне примитивна, поэтому клетки, ее формирующие, не имеют четкого деления на моторные, вставочные и чувствительные. В то же время нужно учитывать, что все же определенное деление клеток нервной системы этого существа существует. Выделяются 2 основных вида нервных клеток - ганглиозные и чувствительные.
Строение этих 2 видов клеток имеет кардинальные отличия. Чувствительные клетки располагаются поперек эпителиального слоя и имеют 1 неподвижный жгутик, усеянный микроскопическими ворсинками. Этот жгутик выходит во внешнюю среду и проводит раздражители, действующие извне. Клетки ганглиозного типа располагаются у самого основания эпителиально-мускульного слоя, поэтому их отростки не могут воспринимать раздражители, действующие извне, но при этом они активно участвуют в сокращении мускулатуры, когда это требуется.
По своему морфологическому составу подавляющее большинство нервных клеток гидры являются биполярными, что обеспечивает им лучшую проводимость и возможность адекватно реагировать на раздражители, воздействующие на тело этого организма из внешней среды.
Несмотря на примитивность строения нервной системы гидры, все же проводимость обеспечивается не только электрическими, но и химическими реакциями. К химическим нейромедиаторам у такого организма, как гидра, относится серотонин, дофамин, гамма-аминокислота, норадриналин, глютамат, глицин, а кроме того больное количество разных видов нейропептидов.
Все эти химические вещества более свойственны сложным животным организмам, но небольшая их часть представлена и у простейших. Несмотря на то что у гидры отсутствует центральная нервная система, все же она способна воспринимать световые раздражители. Сравнительно недавно даже такие организмы, как медузы, считались полностью неспособными различать свет и тьму, но в последствии были обнаружены особые клетки, позволяющие этим существам, дрейфующим по океаническим просторам, различать свет и тьму и выбирать направление движения. Это крайне эффективно, ведь в более поверхностных слоях воды живет большее количество мелких ракообразных и других организмов, которыми питаются медузы.
У гидры имеется схожий механизм распознавания света и тьмы. Распознавать свет гидрам помогает особый чувствительный белок, который также известен, как опсин. Проведение генетического анализа этого белка, извлеченного из тела гидры, позволило выявить ряд схожий черт с аналогичным белком, имеющимся у человека. Подобное исследование показало, что белок опсин у человека и у гидры имеет общее происхождение.
Нервная система гидры довольно эффективна и обеспечивает этому существу лучшие условия для выживания. При минимальном касании к телу гидры, возбуждение которое зарождается в одной точке ее тела, быстро распространяется на другие. Учитывая, что нервный импульс мгновенно распространяется по телу гидры, наблюдается быстрое сокращение кожно-мускульной системы, из-за чего все тело существа быстро укорачивается. Подобная ответная реакция на имеющийся раздражитель извне считается безусловным рефлексом.
Нервные клетки, как и другие ткани тела гидры, отличаются значительной возможностью к регенерации. При разделении гидры на несколько частей каждая из таких половинок в дальнейшем может стать самостоятельным организмом и отрастить утерянные части.
Несмотря на то что гидры, как правило, остаются на одном месте на протяжении длительного времени, все же при необходимости это создание может медленно передвигаться, чтобы найти более удобное место для охоты на свою добычу. Особенности передвижения гидры также во многом обусловлены строением нервной системы этого существа.
Движения гидры. Эпителиально-мускульные клетки эктодермы имеют во-локна, которые могут сокращаться. Если они сокращаются одновремен-но, укорачивается все тело гидры. Если сокращаются волокит в клетках с одной стороны, то в эту сторону гидра наклоняется. Благодаря работе этих волокон движутся щупальца гидры и перемещается все ее тело (рис. 13.4).
Реакции на раздражение гидры. Благодаря нервным клеткам, располо-женным в эктодерме, гидра воспринимает внешние раздражения: свет, прикосновения, некоторые химические вещества. Отростки этих клеток смыкаются между собой, образуя сетку. Так формируется самая простая по строению нервная система, называемая диффузной (рис. 13.5). Больше всего нервных клеток находится возле подошвы и на щупаль-цах. Проявлением работы нервной системы и эпителиально-мышечных клеток является безусловный рефлекс гидры — сгибание щупалец в от-вет на прикосновение.
| Рис. 13.4. Схема движении гидры |
 |
| Рис. 13.5. Нервная система гидры |
Во внешнем слое расположены также стрекательные клетки, содер-жащие капсулы со скрученной тоненькой трубочкой — стрекательной нитью. Наружу из клетки торчит чувствительный волосок. Достаточно его слегка коснуться, как нить выбрасывается из капсулы и вонзается в тело врага или добычи. К нему по стрекательной нити поступает яд, и животное погибает. Больше всего стрекательных клеток расположено в щупальцах.
Регенерация гидры. Маленькие округлые промежуточные клетки эктодер-мы способны превращаться в другие виды клеток. За счет их размноже-ния гидра быстро отстраивает поврежденную часть тела. Способность к регенерации этого животного поражает: когда гидру разделили на 200 частей, из каждой восстановилось целое животное!
Питание гидры. В эндодерме расположены железистые клетки и осна-щенные жгутиками пищеварительные клетки. Железистые клетки по-ставляют в кишечную полость вещества, называемые пищеваритель-ными соками. Эти вещества разрушают добычу, разлагая ее на микроскопические кусочки. С помощью жгутиков пищеварительные клетки подгоняют их к себе и захватывают, образуя псевдоподии. Внутреннюю полость гидры не случайно называют кишечной: в ней начинается переваривание пищи. Но окончательно пища расщепляет-ся в пищеварительных вакуолях пищеварительных клеток. Непе-реваренные остатки пищи удаляются наружу из кишечной полости через рот.
Выделение вредных веществ, образовавшихся в процессе жизнедея-тельности гидры, происходит через эктодерму в воду
Взаимодействие клеток. Среди клеток гидры лишь пищеваритель-ные переваривают пищу, но они обеспечивают питательными вещества-ми не только себя, но и все другие клетки. В свою очередь «соседи» создают наилучшие условия жизни для поставщиков питательных ве-ществ. Вспомните об охоте гидры — теперь вы сможете объяснить, как согласованная работа нервных, жалящих, эпителиально-мышечных и железистых клеток обеспечивает работой пищеварительные клетки. А эти клетки делятся с соседями результатами своего труда. Материал с сайта
Как размножается гидра? При бесполом размножении в результате деления промежуточных клеток образуется почка. Почка растет, на ней появляются щупальца, между ними прорывается рот. На противоположном конце формируется подошва. Маленькая гидра от-деляется от материнского организма, опускается на дно и начинает жить самостоятельно.
Размножается гидра и половым путем. Гидра — гермафродит: в од-них выступах ее эктодермы из промежуточных клеток формируются сперматозоиды, в других — яйцеклетки. Оставив тело гидры, спермато-зоиды по воде следуют к другим особям. Отыскав яйцеклетки, они оплодотворяют их. Образуется зигота, вокруг которой возникает плот-ная оболочка. Это оплодотворенное яйцо остается в теле гидры. Обычно половое размножение происходит осенью. Зимой взрослые гидры по-гибают, а яйца переживают зиму на дне водоема. Весной зигота начина-ет делиться, образуя два слоя клеток. Из них и развивается маленькая гидра.
На этой странице материал по темам:
Статеве розмноження губок
Раздражение и движение доклад биология
Особенности строения и процессов функционирования клеток тела гидры
Особенности процессов жизнедеятельности гидры
Сравнить строение стрекательной клетки гидры и кожицы листа крапивы.
Вопросы по этому материалу:
-
Тело гидры имеет вид продолговатого мешочка, стенки которого состоят из двух слоёв клеток — эктодермы и энтодермы .
Между ними лежит тонкая студенистая неклеточная прослойка — мезоглея , служащая опорой.

Эктодерма формирует покров тела животного и состоит из нескольких видов клеток: эпителиально-мускульные , промежуточные и стрекательные .
Самые многочисленные из них — эпителиально-мускульные.
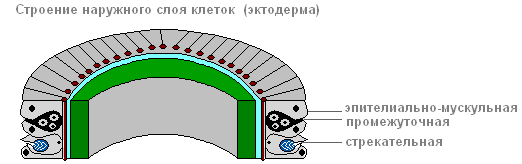
Эктодерма

эпителиально-мускульная клетка
За счёт мускульных волоконец , лежащих в основании каждой клетки, тело гидры может сокращаться, удлиняться и изгибаться.
Между эпителиально-мускульными клетками находятся группы мелких, округлых, с большими ядрами и небольшим количеством цитоплазмы клеток, называемых промежуточными .

При повреждении тела гидры, они начинают усиленно расти и делиться. Они могут превращаться в остальные типы клеток тела гидры, кроме эпителиально-мускульных.
В эктодерме находятся стрекательные клетки , служащие для нападения и защиты. В основном они расположены на щупальцах гидры. Каждая стрекательная клетка содержит овальную капсулу, в которой свёрнута стрекательная нить.
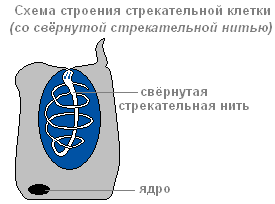
Строение стрекательной клетки со свернутой стрекательной нитью
Если добыча или враг прикоснётся к чувствительному волоску, который расположен снаружи стрекательной клетки, в ответ на раздражение стрекательная нить выбрасывается и вонзается в тело жертвы.
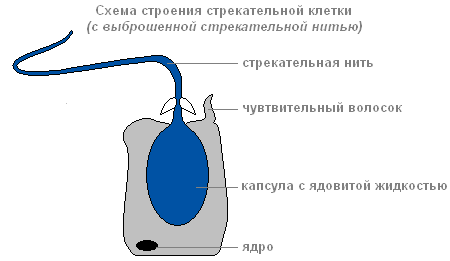
Строение стрекательной клетки с выброшенной стрекательной нитью
По каналу нити в организм жертвы попадает вещество, способное парализовать жертву.
Существует несколько типов стрекательных клеток. Нити одних пробивают кожные покровы животных и вводят в их тело яд. Нити других обвиваются вокруг добычи. Нити третьих — очень клейкие и прилипают к жертве. Обычно гидра «стреляет» несколькими стрекательными клетками. После выстрела стрекательная клетка погибает. Новые стрекательные клетки формируются из промежуточных .
Строение внутреннего слоя клеток
Энтодерма выстилает изнутри всю кишечную полость. В её состав входят пищеварительно-мускульные и железистые клетки.

Энтодерма
Пищеварительная система
Пищеварительно-мускульных клеток больше других. Мускульные волоконца их способны к сокращению. Когда они укорачиваются, тело гидры становится более тонким. Сложные движения (передвижение «кувырканием»), происходит за счёт сокращений мускульных волоконцев клеток эктодермы и энтодермы.
Каждая из пищеварительно-мускульных клеток энтодермы имеет 1-3 жгутика. Колеблющиеся жгутики создают ток воды, которым пищевые частички подгоняются к клеткам. Пищеварительно-мускульные клетки энтодермы способны образовывать ложноножки , захватывать и переваривать в пищеварительных вакуолях мелкие пищевые частицы.
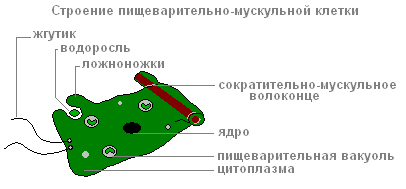
Строение пищеварительно-мускульной клетки
Имеющие в энтодерме железистые клетки выделяют внутрь кишечной полости пищеварительный сок, который разжижает и частично переваривает пищу.

Строение желистой клетки
Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра «надевается» на жертву. Пищеварение начинается в кишечной полости (полостное пищеварение ), заканчивается внутри пищеварительных вакуолей эпителиально-мускульных клеток энтодермы (внутриклеточное пищеварение ). Питательные вещества распределяются по всему телу гидры.
Когда в пищеварительной полости оказываются остатки жертвы, которые невозможно переварить, и отходы клеточного обмена, она сжимается и опорожняется.
Дыхание
Гидра дышит растворённым в воде кислородом. Органов дыхания у неё нет, и она поглощает кислород всей поверхностью тела.
Кровеносная система
Отсутствует.
Выделение
Выделение углекислого газа и других ненужных веществ, образующихся в процессе жизнедеятельности, осуществляется из клеток наружного слоя непосредственно в воду, а из клеток внутреннего слоя — в кишечную полость, затем наружу.
Нервная система
Под кожно-мускульными клетками располагаются клетки звездчатой формы. Это нервные клетки (1). Они соединяются между собой и образуют нервную сеть (2).

Нервная система и раздражимость гидры
Если дотронутся до гидры (2), то в нервных клетках возникает возбуждение (электрические импульсы), которое мгновенно распространяется по всей нервной сети (3) и вызывает сокращение кожно-мускульных клеток и всё тело гидры укорачивается (4). Ответная реакция организма гидры на такое раздражение — безусловный рефлекс .
Половые клетки
С приближением холодов осенью в эктодерме гидры из промежуточных клеток образуются половые клетки.
Различают два вида половых клеток: яйцевые, или женские половые клетки, и сперматозоиды, или мужские половые клетки.
Яйца находятся ближе к основанию гидры, сперматозоиды развиваются в бугорках, расположенных ближе к ротовому отверстию.
Яйцевая клетка гидры похожа на амёбу. Она снабжена ложноножками и быстро растет, поглощая соседние промежуточные клетки.

Строение яйцевой клетки гидры

Строение сперматозоида гидры
Сперматозоиды по внешнему виду напоминают жгутиковых простейших. Они покидают тело гидры и плавают с помощью длинного жгутика.
Оплодотворение. Размножение
Сперматозоид подплывает к гидре с яйцевой клеткой и проникает внутрь нее, причем ядра обеих половых клеток сливаются. После этого ложноножки втягиваются, клетка округляется, на ее поверхности выделяется толстая оболочка — образуется яйцо. Когда гидра погибает и разрушается, яйцо остается живым и падает на дно. С наступлением тёплой погоды живая клетка, находящаяся внутри защитной оболочки, начинает делиться, образующиеся клеточки располагаются в два слоя. Из них развивается маленькая гидра, которая выходит наружу через разрыв оболочки яйца. Таким образом, многоклеточное животное гидра в начале своей жизни состоит всего из одной клетки — яйца. Это говорит о том, что предки гидры были одноклеточными животными.
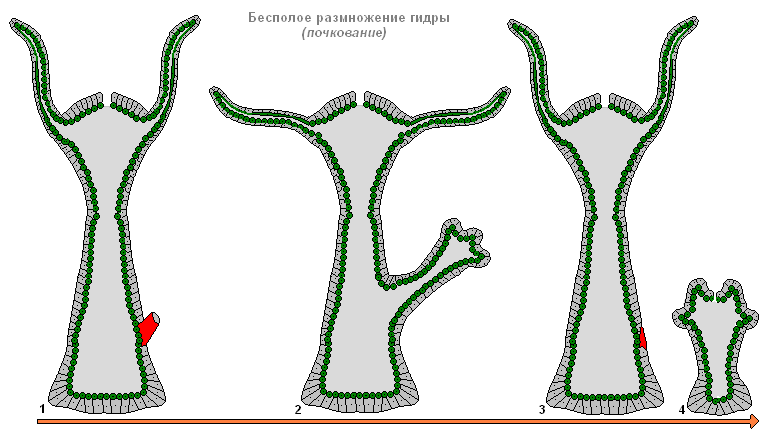
Бесполое размножение гидры
При благоприятных условиях гидра размножается бесполым путём. На теле животного (обычно в нижней трети туловища) образуется почка, она растет, затем формируются щупальца и прорывается рот. Молодая гидра отпочковывается от материнского организма (при этом материнский и дочерний полипы прикрепляются щупальцами к субстрату и тянут в разные стороны) и ведет самостоятельный образ жизни. Осенью гидра переходит к половому размножению. На теле, в эктодерме закладываются гонады - половые железы, а в них из промежуточных клеток развиваются половые клетки. При образовании гонад гидр формируется медузоидный узелок. Это позволяет предполагать, что гонады гидры - сильно упрощенные споросаки, последний этап в ряду преобразования утраченного медузоидного поколения в орган. Большинство видов гидр раздельнополы, реже встречается гермафродитизм. Яйцеклетки гидр быстро растут, фагоцитируя окружающие клетки. Зрелые яйцеклетки достигают диаметра 0,5-1 мм. Оплодотворение происходит в теле гидры: через специальное отверстие в гонаде сперматозоид проникает к яйцеклетке и сливается с ней. Зигота претерпевает полное равномерное дробление, в результате которого образуется целобластула. Затем в результате смешанной деламинации (сочетание иммиграции и деламинации) осуществляется гаструляция. Вокруг зародыша формируется плотная защитная оболочка (эмбриотека) с выростами-шипиками. На стадии гаструлы зародыши впадают в анабиоз. Взрослые гидры погибают, а зародыши опускаются на дно и зимуют. Весной продолжается развитие, в паренхиме энтодермы путем расхождения клеток образуется кишечная полость, затем формируются зачатки щупалец, и из-под оболочки выходит молодая гидра. Таким образом, в отличие от большинства морских гидроидных, у гидры отсутствуют свободноплавающие личинки, развитие у неё прямое.
Регенерация
Гидра обладает очень высокой способностью к регенерации. При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность - рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва - на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела (менее 1/100 объёма), из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса.
Передвижение
В спокойном состоянии щупальца вытягиваются на несколько сантиметров. Животное медленно водит ими из стороны в сторону, подстерегая добычу. При необходимости гидра может медленно передвигаться.
«Шагающий» способ передвижения

«Шагающий» способ передвижения гидры
Изогнув своё тело (1) и прикрепившись щупальцами к поверхности предмета (субстрата), гидра подтягивает к переднему концу тела подошву (2). Затем шагающее движение гидры повторяется (3,4).
«Кувыркающий» способ передвижения

«Кувыркающий» способ передвижения гидры
В другом случае она словно через голову кувыркается, поочерёдно прикрепляясь к предметам то щупальцами, то подошвой (1-5).
Рисунок: Строение пресноводной гидры. Лучевая симетрия гидры
Среда обитания, особенности строения и жизнедеятельности пресноводного полипа гидры
В озерах, речках или прудах с чистой, прозрачной водой на стеблях водных растений встречается маленькое полупрозрачное животное - полип гидра ("полип" означает "многоног"). Это прикрепленное или малоподвижное кишечнополостное животное с многочисленными щупальцами . Тело обыкновенной гидры имеет почти правильную цилиндрическую форму. На одном конце находится рот , окруженный венчиком из 5-12 тонких длинных щупалец, другой конец вытянут в виде стебелька с подошвой на конце. При помощи подошвы гидра прикрепляется к различным подводным предметам. Тело гидры вместе со стебельком обычно длиной до 7 мм, зато щупальца способны вытягиваться на несколько сантиметров.
Лучевая симметрия гидры
Если вдоль тела гидры провести воображаемую ось, то ее щупальца будут расходиться от этой оси во все стороны, как лучи от источника света. Свешиваясь вниз с какого-нибудь водного растения, гидра постоянно покачивается и медленно водит щупальцами, подстерегая добычу. Так как жертва может появиться с любой стороны, лучеобразно расставленные щупальца лучше всего соответствуют такому способу охоты.
Лучевая симметрия характерна, как правило, для животных, ведущих прикпрепленный образ жизни.Кишечная полость гидры
Тело гидры имеет вид мешочка, стенки которого состоят из двух слоев клеток - наружного (эктодермы) и внутреннего (энтодермы). Внутри тела гидры имеется кишечная полость (отсюда и название типа - кишечнополостные).
Наружный слой клеток гидры - эктодерма

Рисунок: строение наружного слоя клеток - эктодермы гидры
Наружный слой клеток гидры называется - эктодерма . Под микроскопом в наружном слое гидры - эктодерме - видно несколько разновидностей клеток. Больше всего здесь кожно-мускулъных. Соприкасаясь боковыми сторонами, эти клетки создают покров гидры. В основании каждой такой клетки есть сократимое мускульное волоконце, играющее важную роль при движении животного. Когда волоконца всех кожно-мускульных клеток сокращаются, тело гидры сжимается. Если сокращаются волоконца только на одной стороне тела, то в эту сторону гидра и нагибается. Благодаря работе мускульных волоконец гидра может медленно передвигаться с места на место, поочередно "ступая" то подошвой, то щупальцами. Такое движение можно сравнить с медленным кувырканием через голову.
В наружном слое расположены и нервные клетки . Они имеют звездообразную форму, так как снабжены длинными отростками.
Отростки соседних нервных клеток соприкасаются между собой и образуютнервное сплетение , охватывающее все тело гидры. Часть отростков подходит к кожно-мускульным клеткам.Раздражимость и рефлексы гидры
Гидра способна ощущать прикосновения, изменение температуры, появление в воде различных растворенных веществ и другие раздражения. От этого ее нервные клетки возбуждаются. Если к гидре прикоснуться тонкой иглой, то возбуждение от раздражения одной из нервных клеток передается по отросткам другим нервным клеткам, а от них - к кожно-мускульным клеткам. Это вызывает сокращение мускульных волоконец, и гидра сжимается в комочек.

Рисунок: раздражимость гидры
На этом примере мы знакомимся со сложным явлением в организме животного - рефлексом . Рефлекс состоит из трех последовательных этапов:восприятия раздражения , передачи возбуждения от этого раздражения по нервным клеткам и ответной реакции организма каким-либо действием. В связи с простотой организации гидры ее рефлексы очень однообразны. В дальнейшем мы ознакомимся с гораздо более сложными рефлексами у более высокоорганизованных животных.
Стрекательные клетки гидры

Рисунок: строкательные или крапивные клетки гидры
Все тело гидры и особенно ее щупальца усажены большим количествомстрекательных , или крапивных клеток. Каждая из этих клеток имеет сложное строение. Кроме цитоплазмы и ядра в ней заключена пузыревидная стрекательная капсула, внутри которой свернута тонкая трубочка -стрекательная нить . Наружу из клетки торчит чувствительный волосок . Как только рачок, малек рыбы или другое маленькое животное коснется чувствительного волоска, стрекательная нить стремительно распрямляется, ее конец выбрасьшается наружу и вонзается в жертву. По каналу, проходящему внутри нити, из стрекательной капсулы в тело добычи попадает яд, вызывающий гибель мелких животных. Как правило, выстреливает сразу много стрекательных клеток. Затем гидра щупальцами подтягивает добычу ко рту и заглатывает. Стрекательные клетки служат гидре также и для защиты. Рыбы и водные насекомые не едят гидр, обжигающих врагов. Яд из капсул по своему действию на организм крупных животных напоминает яд крапивы.
Внутренний слой клеток - энтодерма гидры

Рисунок: строение внутреннего слоя клеток - энтодермы гидры
Внутренний слой клеток - энтодерм а. Клетки внутреннего слоя - энтодермы - имеют сократимые мускульные волоконца, но основная роль этих клеток - переваривание пищи. Они выделяют в кишечную полость пищеварительный сок, под влиянием которого добыча гидры размягчается и распадается на мелкие частицы. Часть клеток внутреннего слоя снабжена несколькими длинными жгутиками (как у жгутиковых простейших). Жгутики находятся в постоянном движении и подгребают частицы к клеткам. Клетки внутреннего слоя способны выпускать ложноножки (как у амебы) и захватывать ими пищу. Дальнейшее пищеварение происходит внутри клетки, в вакуолях (как у простейших). Непереваренные остатки пищи выбрасьшаются наружу через рот.
Особых органов дыхания у гидры нет, растворенный в воде кислород проникает в гидру через всю поверхность ее тела.Регенерация гидры
В наружном слое тела гидры имеются также очень маленькие округлые клетки с крупными ядрами. Эти клетки называют промежуточными . Они играют в жизни гидры очень важную роль. При всяком повреждении тела промежуточные клетки, расположенные вблизи от ран, начинают усиленно расти. Из них образуются кожно-мускульные, нервные и другие клетки, и раненое место быстро зарастает.
Если разрезать гидру поперек, то на одной из ее половинок вырастают щупальца и появляется рот, а на другой возникает стебелек. Получаются две гидры.
Процесс восстановления утраченных или поврежденных частей тела называют регенерацией . У гидры способность к регенерации развита очень сильно.
Регенерация в той или иной степени характерна также для остальных животных и человека. Так, у дождевых червей возможна регенерация целого организма из их частей, у земноводных (лягушки, тритоны) могут восстанавливаться целые конечности, разные части глаза, хвост и внутренние органы. У человека при порезе восстанавливается кожа.Размножение гидры
Бесполое размножение гидры почкованием

Рисунок: бесполое размножение гидры почкованием
Гидра размножается бесполым и половым способами. Летом на теле гидры появляется маленький бугорок — выпячивание стенки ее тела. Бугорок этот растет, вытягивается. На его конце появляются щупальца, а между ними прорывается рот. Так развивается молодая гидра, которая первое время остается соединенной с материнской при помощи стебелька. Внешне все это напоминает развитие побега растения из почки (отсюда и название этого явления - почкование ). Когда маленькая гидра подрастет, она отделяется от материнского организма и начинает жить самостоятельно.
Половое размножение гидры
К осени, с наступлением неблагоприятных условий, гидры гибнут, но перед этим в их теле развиваются половые клетки. Различают два вида половых клеток: яйцевые , или женские, и сперматозоиды , или мужские половые клетки. Сперматозоиды похожи на жгутиковых простейших. Они покидают тело гидры и плавают с помощью длинного жгутика.

Рисунок: половое размножение гидры
Яйцевая клетка гидры похожа на амебу, имеет ложноножки. Сперматозоид подплывает к гидре с яйцевой клеткой и проникает внутрь ее, и ядра обеих половых клеток сливаются. Происходит оплодотворение . После этого ложноножки втягиваются, клетка округляется, на ее поверхности выделяется толстая оболочка — образуется яйцо . В конце осени гидра погибает, а яйцо остается живым и попадает на дно. Весной оплодотворенное яйцо начинает делиться, образующиеся клетки располагаются в два слоя. Из них развивается маленькая гидра, которая с наступлением теплой погоды выходит наружу через разрыв оболочки яйца.
Таким образом, многоклеточное животное гидра в начале своей жизни состоит из одной клетки — яйца.С.М. Никитина, И.А. Ваколюк (Калининградский государственный университет)
Функционирование гормонов как важнейших регуляторов и интеграторов метаболизма и разнообразнейших функций в организме невозможно без существования систем специфической рецепции сигнала и его трансформации в конечный полезный эффект, то есть без гормонкомпетентной системы . Иными словами, наличие реакции на организменном уровне на экзогенные соединения невозможно без наличия циторецепции к этим соединениям и соответственно без существования у этих животных эндогенных соединений, родственных тем, которыми мы воздействуем. Это не противоречит концепции универсальных блоков , когда основные молекулярные структуры в функциональных системах живых организмов обнаруживаются практически в полном наборе уже на самых ранних этапах эволюции, какие только доступны изучению, представлены ограниченным числом молекул и осуществляют одноименные элементарные функции не только у представителей одного царства, например в разных группах млекопитающих или даже в разных типах, но и у представителей различных царств, в том числе у многоклеточных и одноклеточных, у высших эукариот и прокариот.
Однако следует обратить внимание на то, что данные о составе и функциях соединений, выполняющих роль гормонов у позвоночных животных, у представителей таксонов достаточно низкого филогенетического уровня только начинают появляться . Из групп животных низкого филогенетического уровня - гидра как представитель кишечнополостных является наиболее примитивным организмом, обладающим настоящей нервной системой. Нейроны различаются морфологически, химически и, вероятно, функционально. Каждый из них содержит нейросекреторные гранулы . Установлено значительное разнообразие нейрональных фенотипов у гидры. В гипостоме имеются упорядоченно расположенные группы по 6-11 синаптически связанных клеток, которые можно рассматривать как доказательство наличия у гидр примитивных нервных ганглиев. Кроме обеспечения поведенческих реакций, нервная система гидр выполняет роль эндокринной регуляторной системы , обеспечивая контроль метаболизма, размножения, развития. У гидр существует дифференцировка нервных клеток по составу содержащихся в них нейропептидов ). Предполагают , что молекулы окситоцина, вазопрессина, половых стероидов и глюкокортикоидов являются универсальными. Они найдены и у представителей кишечнополостных. Головной и подошвенный активаторы (и ингибиторы) выделены из метаноловых экстрактов тела гидр. Головной активатор, выделенный из актиний, аналогичен по составу и свойствам нейропептиду, обнаруженному в гипоталамусе и кишечнике коровы, крысы, свиньи, человека и в крови последнего. Кроме того, было показано, что и у беспозвоночных и у позвоночных в обеспечении реакции клеток на нейрогормоны участвуют циклические нуклеотиды, то есть механизм действия этих веществ в двух филогенетически различных линиях един.
Целью данного исследования, учитывая вышеизложенное, мы избрали изучение комплексного влияния на пресноводную гидру экзогенных биологически активных (гормональных) соединений.
Материал и методы исследования
Животных для эксперимента собирали в июне-июле 1985-1992 гг. на стационаре (протока реки Немонин, поселок Матросово Полесского района). Адаптация к содержанию в лабораторных условиях - 10-14 суток. Объем материала: тип - Coelenterata; класс - Hydrozoa; вид - Hydra oligactis Pallas; количество - 840. Количество животных отражено в начале эксперимента и не учитывается прирост численности.
В работе были использованы водорастворимые гормональные соединения окситоцинового ряда, передней доли гипофиза с исходной активностью в 1 мл (ип) (гифотоцин - 5ЕД, питуитрин - 5ЕД, маммофизин - 3ЕД, префизон - 25ЕД, гонадотропин - 75ЕД) и стероид - преднизолон - 30 мг, которые у позвоночных обеспечивают трехзвенную эндокринную регуляцию, включающую гипоталамо-гипофизарный комплекс и эпителиальные железы.
В предварительных опытах были использованы концентрации препаратов от 0,00002 до 20 мл ип/л среды содержания животных.
Проводилось три группы исследования:
1-я - определение "+" или "-" реакции во всех принятых нами концентрациях;
2-я - определение диапазона концентраций, обеспечивающих работу в хроническом режиме разной продолжительности;
3-я - хронический эксперимент.
В эксперименте учитывалась активность почкования гидры. Полученные данные подвергались стандартной статистической обработке.
Результаты исследований
При определении "±" реакции гидр в широком диапазоне концентраций соединений были отобраны три (0,1 мл ип/л среды, 0,02 мл ип/л среды и 0,004 мл ип/л среды).
В контрольной группе гидр в течение пяти суток почкование оставалось на уровне 0,0- 0,4 почки /гидру (Ра). В среде минимальной концентрации префизона прирост был 2,2 особи/гидру, питуитрина - 1,9 особи/гидру (достоверность различий с контролем крайне высока - с уровнем значимости 0,01). В средних концентрациях хорошо себя проявили гифотоцин, маммофизин и префизон (1,8-1,9 особей/гидру). Преднизолон в минимальной, и особенно в средней концентрации, вызвал прирост численности 1,1-1,3 особи/гидру, что значительно превышает контроль.
В следующем эксперименте использованы только оптимальные концентрации гормональных соединений. Продолжительность эксперимента - 9 суток. К началу опыта по значению Ра контрольная и экспериментальная группы достоверно не различимы. Через девять суток эксперимента значения Ра достоверно отличалось в опытных группах и контроле с уровнем значимости 0,05 (табл. 1).
Таблица 1
Влияние гормональных препаратов на почкование гидры (Ра) и вероятность достоверности их различий (р)
Среда Ра Изменение р 1 сутки 9 сутки Ра 1 сутки 9 сутки Контроль 1,2±0,8 1,5±0,9 0,3±0,1 - - Гонадотропин 2,1±1,2 5,1±0,3 3,0±0,8 0,71 0,95 Префизон 1,1±0,7 4,9±2,0 3,8±1,3 0,13 0,97 Гифотоцин 1,8±0,8 6,1±2,2 4,3±1,4 0,58 0,99 Питуитрин 0,8±0,5 4,5±2,0 3,7±1,5 0,47 0,98 Маммофизин 1,1±0,3 5,3±2,0 4,2±1,7 0,15 0,99 Преднизолон 1,5±0,4 7,1±2,2 5,6±1,8 0,43 0,99 Как видно из таблицы, наибольшее значение Ра получено при содержании животных в преднизолоне. Все пептидные препараты дают приблизительно сходные значения Ра (в среднем 3,8±0,5). Однако и здесь есть разброс. Наилучший эффект (4,3±1,4) достигается при содержании животных в среде с очищенным экстрактом нейрогипофиза - гифотоцином. Близок к нему по степени воздействия маммофизин. В экспериментальных группах с питуитрином и префизоном значения величины Ра равны 3,7±1,5 и 3,8±1,3 соответственно. Наименьший эффект дает воздействие на гидр гонадотропином. Недостоверные различия в Ра возникают к концу первых суток после помещения гидр в растворы гормональных препаратов. На протяжении девяти суток эксперимента Ра в контроле не изменяется. Начиная с третьих суток Ра во всех экспериментальных группах существенно превышает Ра в контроле. Следует отметить постепенное достоверное увеличение этого показателя у экспериментальных групп к девятым суткам.
Для оценки статистической достоверности оказываемых воздействий сравнивались значения критерия F (отношение средних квадратов), полученные для каждого из двух факторов в отдельности (А - фактор длительности содержания; В - фактор воздействия) и для их взаимодействия (А+В), и табличные значения критерия для двух уровней значимости Р=0,05 и Р=0,01 (табл. 2).
Таблица 2
Результаты дисперсионного анализа влияния гормональных препаратов и длительности содержания на интенсивность бесполого размножения Hydra oligactis
Фак- Фактическое в группах Табличое Р торы Питуитрин Маммофизин Гифотоцин Гонадотропин Префизон Преднизолон 0,05 0,01 А 3,44 1,40 2,27 2,17 3,62 1,30 1,92 2,50 В 8,37 4,04 8,09 4,73 8,26 12,70 4,00 7,08 А+В 1,12 0,96 0,56 0,37 1,07 1,03 1,92 2,50 Как видно из таблицы, Fфакт для фактора воздействия при уровне значимости 0,05 во всех экспериментальных группах больше Fтабл, а при уровне значимости 0,01 такая картина наблюдается в группах с питуитрином, гифотоцином, префизоном и преднизолоном, причем степень воздействия в группе с преднизолоном самая высокая, намного больше, чем в группах с питуитрином, гифотоцином и префизоном, имеющих сходную силу воздействия (значения Fфакт очень близки). Влияние взаимодействия факторов А и В во всех экспериментальных группах не является доказанным.
Для фактора А Fфакт меньше Fтабл (при обоих уровнях значимости) в группах с маммофизином и преднизолоном. В группах с гифотоцином и гонадотропином Fфакт больше Fтабл при Р=0,05, то есть влияние этого фактора не может считаться окончательно доказанным, в отличие от экспериментальных групп с питуитрином и префизоном, где Fфакт больше Fтабл и при Р=0,01 и при Р=0,05.
Все гормональные препараты, кроме гонадотропина, в той или иной степени задерживают начало бесполого размножения. Однако статистически достоверным это оказывается только в группе с префизоном (Р=0,01). Использованные в эксперименте гормональные препараты достоверно не влияют на продолжительность развития единственной почки, изменяют взаимное влияние первой и второй почек: питуитрин, маммофизин, префизон, гонадотропин - при наличии только сформированного головного отдела развивающихся почек; питуитрин, гонадотропин и преднизолон - при наличии хотя бы одного сформированного подошвенного отдела развивающихся почек.
Таким образом, можно считать установленной чувствительность гидр к широкому спектру гормональных соединений позвоночных и предположить, что экзогенные гормональные соединения включаются (как синергисты или антагонисты) в эндокринный регуляторный цикл, присущий самой гидре.
Список литературы
1. Перцева М.Н. Межмолекулярные основы развития гормонкомпетентности. Л.: Наука, 1989.
2. Богута К.К. Некоторые морфологические принципы формирования низкоорганизованных нервных систем в онто- и филогенезе // Успехи современной биологии. М.: Наука, 1986. Т. 101. Вып. 3.
3. Иванова-Казас А.А. Бесполое размножение животных. Л., 1971.
4. Наследов Г.А. Многовариантность осуществления элементарных функциональных задач и упрощение системы молекулярных взаимодействий как закономерность функциональной эволюции // Журнал эволюционной биохимии и физиологии. 1991. Т. 27. № 5.
5. Наточин Ю.В., Бройнлих Х. Использование методов токсикологии в изучении проблемы эволюции функций почки // Журнал эволюционной биохимии и физиологии. 1991. Т. 27. № 5.
6. Никитина С.М. Стероидные гомоны у беспозвоночных животных: Монография. Л.: Изд-во ЛГУ, 1987.
7. Афонькин С.Ю. Межклеточное самораспознование у простейших // Итоги науки и техники. М., 1991. Т. 9.
8. Проссер Л. Сравнительная физиология животных. М.: Мир, 1977. Т. 3.
9. Резников К.Ю., Назаревская Г.Д. Стратегия развития нервной системы в онто- и филогенезе. Гидра // Успехи современной биологии. М.: Наука, 1988. Т. 106. Вып.2 (5).
10. Шейман И.М., Балобанова Э.Ф., Пептидные гормоны беспозвоночных // Успехи современной биологии. М.: Наука, 1986. Т. 101. Вып. 2.
11. Этингоф Р.Н. Изучение молекулярной структуры нейрорецепторов. Методические подходы, эволюционные аспекты // Журнал эволюционной биохимии и физиологии. 1991. Т. 27. № 5.
12. Highnam K.C., Hill L. The comparative Endocrinology of the Invertebrates // Edward Arnold, 1





