Hemo - sang et armure. globus - boule) est une molécule protéique complexe située à l'intérieur des globules rouges - érythrocytes (chez l'homme et les vertébrés). L'hémoglobine représente environ 98 % de la masse de toutes les protéines des globules rouges.
Hémoglobine(du grec ancien hémo - sang et lat. globus - boule) est une molécule protéique complexe située à l'intérieur des globules rouges - les érythrocytes (chez l'homme et les vertébrés). L'hémoglobine représente environ 98 % de la masse de toutes les protéines des globules rouges. En raison de sa structure, l'hémoglobine participe au transfert de l'oxygène des poumons vers les tissus, et au retour du monoxyde de carbone.
La structure de l'hémoglobine
L'hémoglobine est constituée de deux chaînes de globine de type alpha et de deux chaînes de l'autre type (bêta, gamma ou sigma), reliées à quatre molécules d'hème, qui contiennent du fer. La structure de l’hémoglobine s’écrit avec les lettres de l’alphabet grec : α2γ2.
Échange d'hémoglobine
L'hémoglobine est formée par les globules rouges de la moelle osseuse rouge et circule avec les cellules tout au long de leur vie, soit 120 jours. Lorsque les vieilles cellules sont éliminées par la rate, les composants de l’hémoglobine sont éliminés du corps ou relâchés dans la circulation sanguine pour être incorporés dans de nouvelles cellules.
Types d'hémoglobine
Les types normaux d'hémoglobine comprennent l'hémoglobine A ou HbA (d'adulte à adulte), ayant la structure α2β2, l'HbA2 (hémoglobine mineure d'un adulte, ayant la structure α2σ2 et l'hémoglobine fœtale (HbF, α2γ2. Hémoglobine F - hémoglobine fœtale. Remplacement par l'adulte l'hémoglobine apparaît complètement vers 4 à 6 mois (le taux d'hémoglobine fœtale à cet âge est inférieur à 1%). L'hémoglobine fœtale se forme 2 semaines après la fécondation, plus tard, après la formation du foie fœtal, elle est remplacée par l'hémoglobine fœtale.
Il existe plus de 300 hémoglobines anormales, elles portent le nom du lieu de découverte.
Fonction d'hémoglobine
La fonction principale de l’hémoglobine est de transporter l’oxygène des poumons vers les tissus et de renvoyer le dioxyde de carbone.
Formes d'hémoglobine
- Oxyhémoglobine- combinaison d'hémoglobine et d'oxygène. L'oxyhémoglobine prédomine dans le sang artériel allant des poumons aux tissus. En raison de la teneur en oxyhémoglobine, le sang artériel a une couleur écarlate.
- Hémoglobine réduite ou désoxyhémoglobine(HbH) - hémoglobine qui donne de l'oxygène aux tissus
- Carboxyhémoglobine- combinaison d'hémoglobine et de dioxyde de carbone. On le trouve dans le sang veineux et lui donne une couleur cerise foncée.
 Effet Bohr
Effet Bohr
L'effet a été décrit par le physiologiste danois Christian Bohr http://en.wikipedia.org/wiki/Christian_Bohr (père du célèbre physicien Niels Bohr).
Christian Bohr précise qu'avec une plus grande acidité (un pH plus faible, par exemple dans les tissus), l'hémoglobine se liera moins à l'oxygène, ce qui lui permettra d'être libéré.
Dans les poumons, dans des conditions d'excès d'oxygène, il se combine à l'hémoglobine des globules rouges. Les globules rouges transportent l'oxygène dans le sang vers tous les organes et tissus. Des réactions d'oxydation se produisent dans les tissus du corps avec la participation de l'oxygène entrant. À la suite de ces réactions, des produits de décomposition se forment, notamment du dioxyde de carbone. Le dioxyde de carbone des tissus est transféré aux globules rouges, ce qui diminue l'affinité pour l'oxygène et libère de l'oxygène dans les tissus.
Effet Bohr est d'une grande importance pour le fonctionnement du corps. Après tout, si les cellules travaillent intensément et libèrent plus de CO2, les globules rouges peuvent leur fournir plus d’oxygène, évitant ainsi le « manque » d’oxygène. Ces cellules peuvent donc continuer à fonctionner à un rythme élevé.
 Quel est le taux d’hémoglobine normal ?
Quel est le taux d’hémoglobine normal ?
Chaque millilitre de sang contient environ 150 mg d'hémoglobine ! Les niveaux d'hémoglobine changent avec l'âge et dépendent du sexe. Ainsi, l'hémoglobine chez les nouveau-nés est nettement plus élevée que chez les adultes et chez les hommes, elle est plus élevée que chez les femmes.
Qu'est-ce qui affecte les niveaux d'hémoglobine ?
Certaines autres conditions affectent également les niveaux d'hémoglobine, comme l'exposition à l'altitude, le tabagisme et la grossesse.
Maladies associées à des modifications de la quantité ou de la structure de l'hémoglobine
- Une augmentation des taux d'hémoglobine est observée avec l'érythrocytose et la déshydratation.
- Une diminution des taux d'hémoglobine est observée dans diverses anémies.
- En cas d'intoxication au monoxyde de carbone, il se forme de la carbhémoglobine (à ne pas confondre avec la carboxyhémoglobine !), qui ne peut pas fixer l'oxygène.
- Sous l'influence de certaines substances, de la méthémoglobine se forme.
- Un changement dans la structure de l’hémoglobine est appelé hémoglobinopathie. Les maladies les plus connues et les plus courantes de ce groupe sont la drépanocytose, la bêta-thalassémie et la persistance de l'hémoglobine fœtale. Voir les hémoglobinopathies sur le site de l'Organisation Mondiale de la Santé
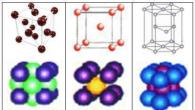
La molécule d'hémoglobine contient 4 groupes hème identiques. L'hème est une porphyrine contenant un ion Fe 2+ situé au centre. C'est un dérivé de la porphine, qui est un système condensé de 4 pyrroles reliés par des ponts méthine (-CH=). Selon la structure des substituants de la porphine, on distingue plusieurs types d'hèmes.
L'hème IX est le type d'hème le plus courant. Le dérivé de porphine qu'il contient est la protoporphyrine IX (porphine de l'acide 1,3,5,8 - tétraméthyl-2,4 - divinyl - 6, 7 - dipropionique) ;
hème a (formylporphyrine). L'hème a, au lieu d'un groupe méthyle, contient un résidu formyle en huitième position (-CHO) et au lieu d'un groupe vinyle (en deuxième position) une chaîne isoprénoïde. L'hème a fait partie de la cytochrome oxydase ;
hème c, dans lequel les résidus de cystéine sont associés à des groupes vinyle (-CH=CH 2) en positions 2 et 4. Une partie du cytochrome C ;
l'hème est une dihydroporphyrine de fer 4.
L'hème est un groupe prothétique non seulement de l'hémoglobine et de ses dérivés, mais également de la myoglobine, de la catalase, de la peroxydase, des cytochromes et de l'enzyme tryptophane pyrollase, qui catalyse l'oxydation du troptophane en formylkynurénine.
Le numéro de coordination des atomes de fer est 6. Dans l'hème, le fer est lié par deux liaisons covalentes aux atomes d'azote de deux cycles pyrrole et par deux liaisons de coordination aux atomes d'azote des cycles pyrrole restants. Les cinquième et sixième liaisons de coordination du fer sont réparties différemment selon la molécule protéique qui contient de l'hème et selon ses fonctions. Par exemple, dans les cytochromes 5 et 6, les liaisons de coordination du fer sont liées aux résidus histidine et méthionine. Cet arrangement de l'hème dans les cytochromes est nécessaire pour remplir leur fonction spécifique : le transfert d'électrons dans la chaîne respiratoire. Transitions Fe 3+ + e= Fe 2+ ; Fe 2+ -е= Fe 3+ crée la possibilité de transférer des électrons d'un cytochrome à un autre.
Examinons de plus près l'emplacement de l'hème dans l'hémoglobine (myoglobine). L'hème est situé dans la fente entre les hélices E et F ; ses groupes propionates polaires sont orientés vers la surface du globule, et le reste est situé à l'intérieur de la structure et est entouré de résidus apolaires, à l'exception de His F8 et His F7. La cinquième position de coordination de l'atome de fer est occupée par l'atome d'azote de l'hétérocycle de l'histidine proximale His F8. L'histidine distale (His F7) est située de l'autre côté du cycle hème, presque en face de His F8, mais la sixième position de coordination de l'atome de fer reste vacante. Parmi les deux liaisons de coordination inutilisées, l'une va se connecter à la protéine et la seconde va se connecter à divers ligands (physiologiques - oxygène, eau et étrangers - dioxyde de carbone, cyanure, etc.).
Dérivés de l'hémoglobine
L'hémoglobine interagit avec divers ligands ; la sixième liaison de coordination du fer dans l'hème est destinée à cela. Les dérivés de l'hémoglobine comprennent :
oxyhémoglobine HbO 2 – un composé d'oxygène moléculaire avec l'hémoglobine. Pour souligner le fait que la valence du fer ne change pas au cours de cette liaison, la réaction est appelée oxygénation plutôt qu'oxydation ; le processus inverse est appelé désoxygénation.
Lorsqu’ils veulent préciser spécifiquement que l’hémoglobine n’est pas associée à l’oxygène, on l’appelle désoxyhémoglobine ;
carboxyhémoglobine HbCO.
cyanméthémoglobine (HbСN) – la méthémoglobine a également un effet positif. Il lie le CN - pour former de la cyanméthémoglobine et sauve le corps des effets mortels du cyanure. Par conséquent, les formateurs de méthémoglobine (le même nitrite de Na) sont utilisés pour traiter l'empoisonnement au cyanure ;
la carbhémoglobine se forme lorsque l'hémoglobine se lie au CO 2 .
Cependant, le CO 2 ne s'attache pas à l'hème, mais aux groupes NH 2 de la globine :
HbNH 2 + CO 2 = HbNHCOO - + H +
De plus, la désoxyhémoglobine fixe plus de CO 2 que l'oxyhémoglobine. La formation de carbhémoglobine est utilisée pour éliminer le CO 2 des tissus vers les poumons. 10 à 15 % de CO 2 sont ainsi éliminés.
Grâce à la sixième liaison de coordination, une molécule d'oxygène s'attache à l'atome de fer pour former de l'oxyhémoglobine. Les anneaux pyrrole de l’hème sont situés dans le même plan, tandis que l’atome de fer dépasse quelque peu de ce plan. L'ajout d'oxygène « redresse » la molécule d'hème : le fer se déplace dans le plan des anneaux pyrrole de 0,06 nm, puisque le diamètre de la sphère de coordination de l'atome de fer diminue. L'hémoglobine lie 4 molécules d'oxygène (une molécule par hème dans chaque sous-unité). L'oxygénation s'accompagne de changements conformationnels importants de l'hémoglobine. En se déplaçant dans le plan des anneaux pyrrole, Fe, connecté en 5ème position de coordination au résidu HisF8, « tire » la chaîne peptidique vers elle. Il y a un changement dans la conformation de cette chaîne et des autres chaînes polypeptidiques qui lui sont associées, puisqu'un protomère est relié par de nombreuses liaisons à d'autres protomères. Ce phénomène est appelé coopérativité des changements dans la conformation des protomères. Les changements conformationnels sont tels que la liaison initiale de l'O 2 à une sous-unité accélère la liaison des molécules d'oxygène aux sous-unités restantes. Ce phénomène est connu sous le nom d’effet coopératif positif homotrope (homotrope car seul l’oxygène est impliqué). C'est ce qui détermine le caractère sigmoïde de la courbe de saturation en oxygène de l'hémoglobine. La quatrième molécule d'oxygène s'attache à l'hémoglobine 300 fois plus facilement que la première molécule. Pour avoir une idée plus précise de ce mécanisme, il convient de considérer la structure de l'hémoglobine sous la forme de deux hétérodimères formés de et - sous-unités : 1 1 et 2 2. Un léger déplacement de l'atome de fer conduit au fait qu'une paire de sous-unités / tourne par rapport à l'autre paire / . Dans ce cas, les liaisons non covalentes entre sous-unités provoquées par les interactions électrostatiques sont détruites. Un ensemble de liaisons entre dimères est remplacé par un autre et leur rotation relative se produit.
La structure quaternaire de l'hémoglobine partiellement oxygénée est décrite comme un état T (de l'anglais Taut - tension), l'hémoglobine entièrement oxygénée (HbO 2) correspond à un état R - (détendu - relaxation). La condition est caractérisée par une affinité plus faible pour l'oxygène ; la probabilité de transition de la forme T à la forme R augmente à mesure que chacun des 4 hémogroupes est oxygéné séquentiellement. Les ponts salins (liaisons non covalentes) sont détruits à mesure que de l'oxygène est ajouté, augmentant ainsi la probabilité d'une transition de la forme T à la forme R (état de haute affinité).
L'hémoglobine fait partie du groupe des protéines hémoprotéines, qui sont elles-mêmes un sous-type de chromoprotéines et sont divisées en non enzymatique protéines (hémoglobine, myoglobine) et enzymes (cytochromes, catalase, peroxydase). Leur partie non protéique est l'hème - une structure qui comprend un cycle porphyrine (constitué de 4 cycles pyrrole) et un ion Fe 2+. Le fer se lie à l’anneau porphyrine avec deux liaisons de coordination et deux liaisons covalentes.
La structure de l'hémoglobine
La structure de l'hémoglobine A
L'hémoglobine est une protéine constituée de 4 sous-unités protéiques contenant de l'hème. Les protomères sont reliés les uns aux autres par des liaisons hydrophobes, ioniques et hydrogène, et n’interagissent pas arbitrairement, mais via une zone spécifique : la surface de contact. Ce processus est très spécifique, le contact se produit simultanément en des dizaines de points selon le principe de complémentarité. L'interaction est réalisée par des groupes de charges opposées, des régions hydrophobes et des irrégularités à la surface de la protéine.
Les sous-unités protéiques de l'hémoglobine normale peuvent être représentées par différents types de chaînes polypeptidiques : α, β, γ, δ, ε, ξ (respectivement grec - alpha, bêta, gamma, delta, epsilon, xi). La molécule d'hémoglobine contient : deux chaînes deux différents types.
L'hème se combine avec la sous-unité protéique, d'abord par l'intermédiaire du résidu histidine lien de coordination du fer, deuxièmement, à travers liaisons hydrophobes anneaux pyrrole et acides aminés hydrophobes. L'hème se trouve en quelque sorte « dans une poche » de sa chaîne et un protomère contenant de l'hème se forme.
Formes normales d'hémoglobine
Il existe plusieurs variantes normales de l'hémoglobine :
- HbР – hémoglobine primitive, contient des chaînes 2ξ et 2ε, se trouve dans l'embryon entre 7 et 12 semaines de vie,
- HbF – hémoglobine fœtale, contient des chaînes 2α et 2γ, apparaît après 12 semaines de développement intra-utérin et est la principale après 3 mois,
- HbA – hémoglobine adulte, la proportion est de 98 %, contient des chaînes 2α et 2β, apparaît chez le fœtus après 3 mois de vie et représente à la naissance 80 % de toute l'hémoglobine,
- HbA 2 – hémoglobine des adultes, la proportion est de 2%, contient des chaînes 2α et 2δ,
- HbO 2 - oxyhémoglobine, est formée par la liaison de l'oxygène dans les poumons, dans les veines pulmonaires, elle représente 94 à 98 % de la quantité totale d'hémoglobine,
- HbCO 2 – carbohémoglobine, est formée par la liaison du dioxyde de carbone dans les tissus ; dans le sang veineux, elle représente 15 à 20 % de la quantité totale d'hémoglobine.
Formes pathologiques de l'hémoglobine
HbS – hémoglobine drépanocytaire.
MetHb – méthémoglobine, une forme d'hémoglobine qui comprend un ion ferrique au lieu d'un ion ferreux. Cette forme se forme spontanément, lors de l'interaction d'une molécule d'O 2 et de l'hème Fe 2+, mais généralement le pouvoir enzymatique de la cellule suffit à la restituer. Lors de l'utilisation de sulfamides, de la consommation de nitrite et de nitrates de sodium dans les produits alimentaires et en cas d'insuffisance d'acide ascorbique, la transition de Fe 2+ en Fe3+ est accélérée. La metHb qui en résulte n’est pas capable de lier l’oxygène et une hypoxie tissulaire se produit. Pour réduire Fe3+ en Fe2+, la clinique utilise de l'acide ascorbique et du bleu de méthylène.
L'hémoglobine fait partie des globules rouges et remplit la majeure partie de leur espace intracellulaire. La fonction principale de l'hémoglobine est liée au transport des gaz (oxygène et dioxyde de carbone) dans le sang humain. De plus, l'hémoglobine participe au maintien de l'équilibre acido-basique dans le corps humain et animal, formant le système tampon d'hémoglobine le plus puissant du sang.
Actuellement, la structure et les propriétés de l'hémoglobine sont assez bien étudiées. Chez un adulte, on distingue les types physiologiques d'hémoglobine suivants dans le sang :
1. Hémoglobine A 1 (HbA 1 – de l'anglais. adulte– adulte), dont la teneur est de 96 % de l’hémoglobine totale (Hb).
2. Hémoglobine A 2 (HbA 2) - la teneur peut atteindre 2,5 %.
3. Hémoglobine fœtale (HbF de l'anglais. fœtus- fruits) est de 1,5 à 2 %.
L'HbF est la principale hémoglobine du fœtus et des nouveau-nés, puisque sa teneur chez les nouveau-nés atteint jusqu'à 80 %, mais au cours des trois premiers mois après la naissance, elle est presque entièrement remplacée par l'HbA.
Sur la fig. La figure 1 montre schématiquement la structure de la molécule d'hémoglobine.
Riz. 1. Modèle de la molécule d'hémoglobine (HbA 1) (Lanzug avec chaîne ukrainienne).
La molécule d'hémoglobine adulte HbA 1 est constituée de quatre chaînes polypeptidiques, chacune étant associée à un hème. La partie protéique de la molécule d'hémoglobine est appelée « globine ».
L'HbA 1 comprend les chaînes 2a et 2b, qui sont les produits de l'expression de deux gènes différents et ont donc des structures primaires différentes. La chaîne a en contient 141 et la chaîne b contient 146 résidus d’acides aminés. Les sous-unités d'hémoglobine, contenant chacune une chaîne polypeptidique et un hème, ressemblent à la structure de la molécule de myoglobine dans leur conformation (Fig. 7). Schématiquement, l'hémoglobine A 1 s'écrit comme suit : HbA 1 = α 2 β 2. Dans l'hémoglobine A 2, au lieu des sous-unités β, il y a des sous-unités δ : HbA 2 = α 2 δ 2, et dans l'hémoglobine fœtale, il y a des sous-unités γ, c'est-à-dire HbF = α 2 γ 2.
Lors de la formation de la structure quaternaire de l'hémoglobine, de nombreuses liaisons non covalentes apparaissent entre les chaînes polypeptidiques individuelles de la globine. Le plus grand nombre d'entre elles est formée entre différents types de chaînes (a - b, α - δ, α - γ). Il s'agit d'interactions majoritairement hydrophobes qui se produisent entre les radicaux de certains acides aminés (leucine, valine, phénylalanine...). L'étude de l'organisation structurale des sous-unités de la molécule d'hémoglobine en protéine oligomère a été réalisée à l'aide d'une solution d'urée 8 M ou avec des changements brusques de pH. Dans ce cas, la molécule d’hémoglobine se dissocie de manière réversible en deux chaînes α et deux chaînes β. Cette dissociation est provoquée par la rupture des liaisons hydrogène. Après élimination de l'urée, une association automatique de la molécule d'hémoglobine d'origine se produit (Fig. 2)

Riz. 2. Dissociation réversible de la molécule d'hémoglobine sous l'action d'une solution d'urée 8 M.
Le composant non protéique de l’hémoglobine est l’hème. La base de la structure de l'hème est la protoporphyrine. La protoporphyrine est constituée de quatre anneaux pyrrole reliés par des ponts α-méthine (–CH=). Selon la nature des groupes situés dans les radicaux latéraux, les porphyrines possèdent un grand nombre d'isomères. Parmi les 15 isomères possibles des protoporphyrines, le plus répandu dans les objets biologiques est la protoporphyrine IX. Il contient 4 groupes méthyle, 2 vinyles et 2 groupes propionyle en positions latérales (Fig. 3 A). Le complexe chélaté de la protoporphyrine IX avec Fe 2+ est appelé protohème IX ou hème.
Le cation fer inclus dans la structure hème forme deux liaisons covalentes et deux liaisons de coordination avec les atomes d'azote des cycles pyrrole dans le plan du complexe protoporphyrine. De plus, il participe à la formation de deux autres liaisons de coordination, situées perpendiculairement au plan du complexe protoporphyrine (Fig. 3 B).
Le système circulatoire remplit une fonction de transport dans le corps de tous les animaux à sang chaud, fournissant des nutriments et de l'oxygène aux tissus. Le transport de l'oxygène et du dioxyde de carbone s'effectue grâce aux globules rouges, qui contiennent une substance importante - l'hémoglobine. Dans cet article, nous examinerons les types et les composés de l'hémoglobine.
Qu'est-ce que l'hémoglobine
L'hémoglobine est un composant des globules rouges qui appartient au groupe des protéines. Se compose de 96 % de substance protéique globine et de 4 % de substance avec un atome - hème. Une cellule érythrocytaire contient environ 280 millions de molécules qui donnent la couleur rouge au sang.
La principale propriété de l'hémoglobine est la capacité du fer à attacher et à séparer les gaz, formant ainsi le mouvement de l'oxygène des poumons vers les tissus et du dioxyde de carbone des tissus vers les poumons. Ainsi, son rôle dans le processus d’échange gazeux dans le corps est irremplaçable.
Structure et types d'hémoglobine dans le sang humain
À différents stades de développement du corps humain, la composition de l'hémoglobine diffère par la structure des chaînes polypeptidiques. Selon les chaînes polypeptidiques que contient la structure de l'hémoglobine, les types d'hémoglobine chez l'homme sont les suivants :

Types pathologiques d'hémoglobine
Dans certains cas, sous l'influence de défauts génétiques, une synthèse anormale des cellules d'hémoglobine se produit. Les types pathologiques d'hémoglobine diffèrent des types physiologiques par la composition des liaisons polypeptidiques, ou plus précisément par leur mutation.
À la suite d'une mutation de l'ADN, la synthèse des composants érythrocytaires s'effectue non pas avec de la glutamine, mais avec l'acide aminé valine. Ce remplacement de « trame » conduit à la formation d’une structure protéique de type 2 avec une région « collante » à la surface qui est capable d’attribuer des structures à son propre type. Ainsi, il se produit une polymérisation des molécules d'HbS et, par conséquent, une sédimentation de globules rouges lourds et mal transportés dans les vaisseaux sanguins. Cette déviation est appelée

Valeurs normales de la teneur en hémoglobine dans le sang, non considérées comme un écart pathologique :
- Pour les hommes – 130-150 g/l.
- Pour les femmes - 120-140 g/l.
- Chez les enfants de moins d'un an, 100-140 g/l, et au cours du premier mois, ces valeurs peuvent atteindre jusqu'à 220 g/l en raison de l'augmentation de la concentration d'hémoglobine fœtale. Chez les enfants de un à 6 ans - 110-145 g/l et à partir de 6 ans - 115-150 g/l, quel que soit le sexe de l'enfant.
- Pendant la grossesse, on observe une diminution de la concentration d'HbA jusqu'à 110 g/l, ce qui n'est cependant pas considéré comme une anémie.
- Chez les personnes âgées, la norme est considérée comme une tendance à diminuer de 5 unités par rapport à la norme indiquée, en fonction du sexe du patient.
La composition du sang, qui contient simultanément différents types d'hémoglobine, diffère également. Par exemple, chez un adulte, le ratio naturel est de 99 % d'HbA et jusqu'à 1 % d'HbF. Chez les enfants de moins d'un an, le pourcentage d'HbF est nettement plus élevé que chez les adultes, ce qui s'explique par la dégradation progressive de la forme initialement existante d'hémoglobine fœtale.
Formes physiologiques
Étant donné que le pigment rouge respiratoire est continuellement impliqué dans les processus d'échange gazeux dans le corps, sa principale propriété est la capacité de former des composés avec des molécules de divers gaz. À la suite de telles réactions, des types physiologiques d’hémoglobine sont créés, considérés comme normaux.

Connexions pathologiques
Les globules rouges peuvent fixer non seulement les gaz impliqués dans le processus respiratoire, mais également d'autres, formant des types pathologiques d'hémoglobine qui présentent un danger pour la santé humaine, voire la vie. Ces composés ont un faible degré de décomposition et entraînent donc un manque d'oxygène dans les tissus et de graves perturbations du processus respiratoire.

Diagnostic de l'hémoglobine
Pour identifier la concentration de structures respiratoires de globine dans le sang humain, des analyses qualitatives et quantitatives sont effectuées. L'hémoglobine est également examinée pour la quantité d'ions fer qu'elle contient.

Aujourd'hui, la principale méthode quantitative permettant de déterminer la concentration d'hémoglobine est l'analyse colorimétrique. Il s'agit d'une étude de la saturation des couleurs d'un matériau biologique lorsqu'un réactif spécial y est ajouté.
Les méthodes qualitatives comprennent des analyses de sang pour déterminer le rapport entre les types d'HbA et d'HbF. Cela comprend également la détermination de la quantité de molécules d'hémoglobine glyquée (composés avec des carbones) dans le sang - la méthode est utilisée pour diagnostiquer le diabète sucré.
Écart de la concentration d'hémoglobine par rapport à la normale
Le solde HbA peut varier à la fois en dessous et au-dessus de la normale. Dans tous les cas, cela entraîne des conséquences négatives. Lorsque l’HbA descend en dessous de la norme établie, un syndrome pathologique apparaît, appelé « anémie ferriprive ». Exprimé par la léthargie, la perte de force, l'inattention. Il a un effet négatif sur le système nerveux et est particulièrement dangereux pendant l'enfance, car il provoque souvent un retard du développement psychomoteur.

L'augmentation du taux d'hémoglobine n'est pas une maladie distincte ; il s'agit plutôt d'un syndrome indiquant diverses pathologies, telles que le diabète sucré, l'insuffisance pulmonaire, les maladies cardiaques, les maladies rénales, l'excès d'acide folique ou de vitamines B, l'oncologie, etc.